PISUM GENETICS
2011-VOLUME 43
RESEARCH PAPERS
Use of Supernodulating Mutants in Pea Breeding
Sidorova, K.K. Russian Academy of Sciences - Siberian Branch, Novosibirsk, Russia
Our genetic studies of symbiotic pea mutants have shown that the Pisum genome has multiple genes
controlling the following traits: (1) capability of symbiosis with Rhizobium nodule bacteria; (2) the
number and efficiency of root nodules; (3) nitrogen fixation rate intensity, assessed from nitrogenase
activity; (4) duration of active nitrogen fixation; and (5) root biomass accumulation. We conclude that
the macrosymbiont plays the key role in the genetic control of nitrogen fixation in nodules and that
breeding of legume macrosymbionts for greater efficiency of their symbiosis with rhizobia is promising
given that rhizobia are present in any substrate or soil, in the field or in a greenhouse.
Supernodulating mutants deserve special attention of breeders as a means for increasing nodulation and
nitrogen fixation. However, these mutants suffer from low productivity. The first attempts to involve
supernodulating mutants in breeding
were done with soybean, despite
being unsuccessful (1). To test the
feasibility of utilizing
supernodulating mutants in legume
breeding as tools for intense nitrogen
fixation, we performed large-scale
field and greenhouse experiments
with pea and investigated the
expression of the nod4 and nod3
supernodulating genes against
various genetic backgrounds (2). The
results were used in the development
of a breeding program for improving
the efficiency of the legume-
rhizobium symbiosis (Figure 1).
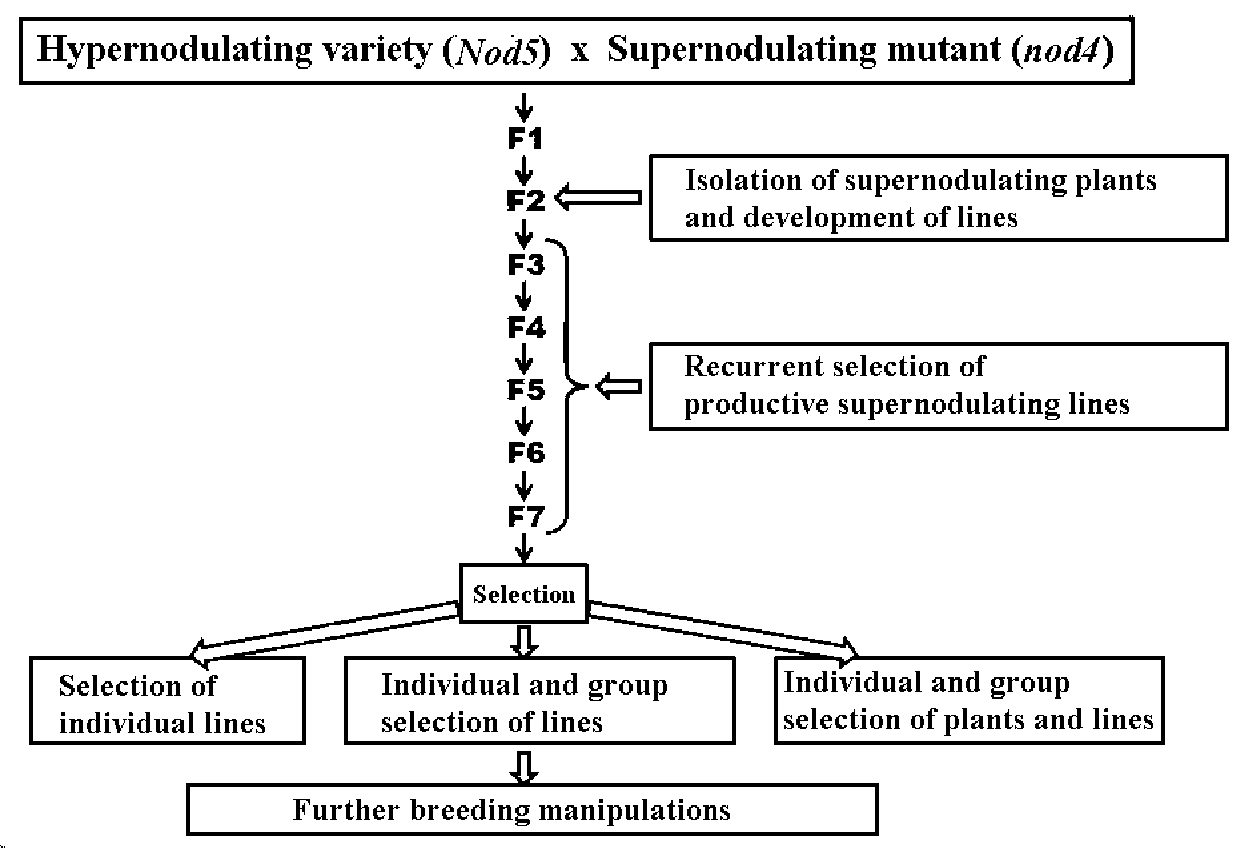
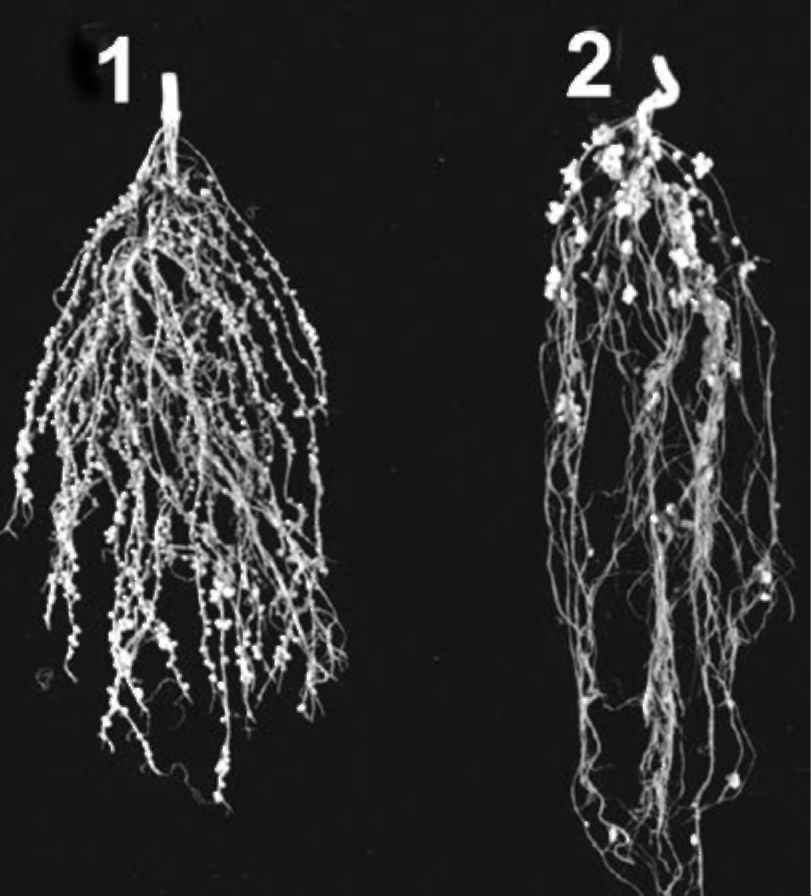
Figure 1. Breeding program for increasing nitrogen fixation in pea Pisum sativum L.
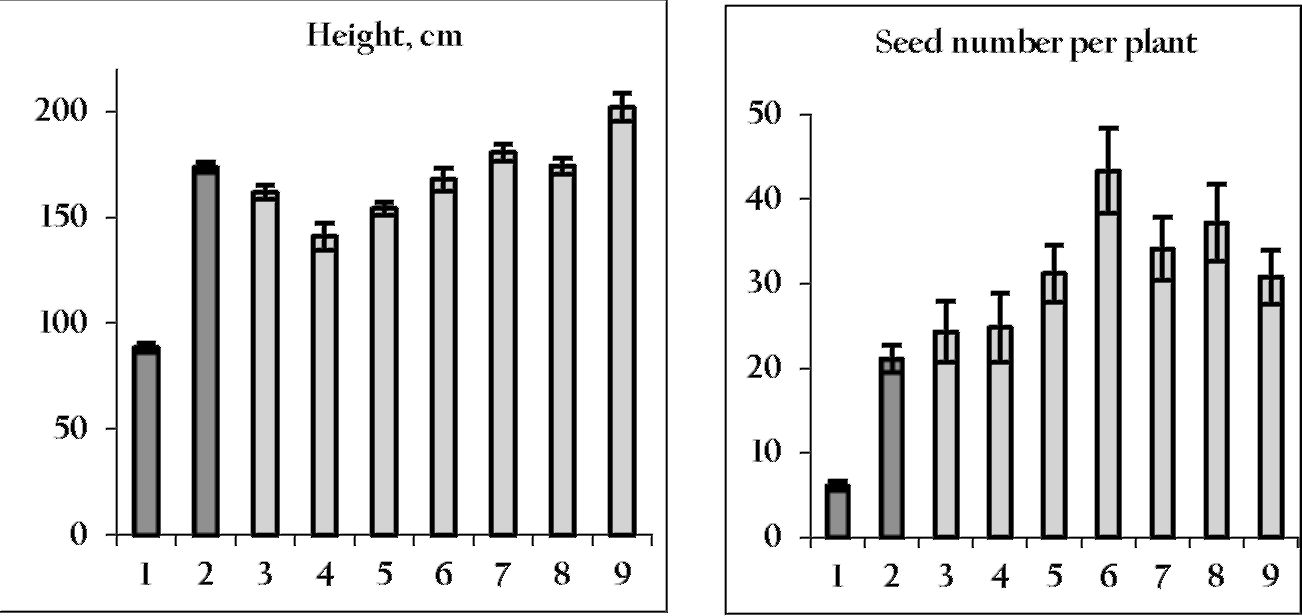
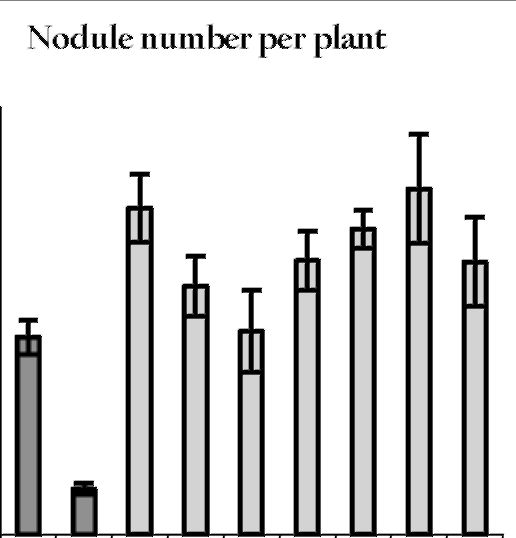
it is important to choose a proper variety, which
should be highly productive and hypernodulating.
Hypernodulation is characterized by large nodules,
forming mainly in the upper and middle portions of
roots. The nodules are considerably fewer in number,
but larger than in supernodulating forms (Figure 2).
Hypernodulation is governed by the dominant allele at
the Nod5 locus located on linkage group (LG) III (3, 4).
The pea germplasm collection at Novosibirsk includes
pea mutants and varieties with hypernodulation for
Nod5. The supernodulation in mutant K301, used in our
study, has the recessive allele at the nod4 locus located
in LG V (5).
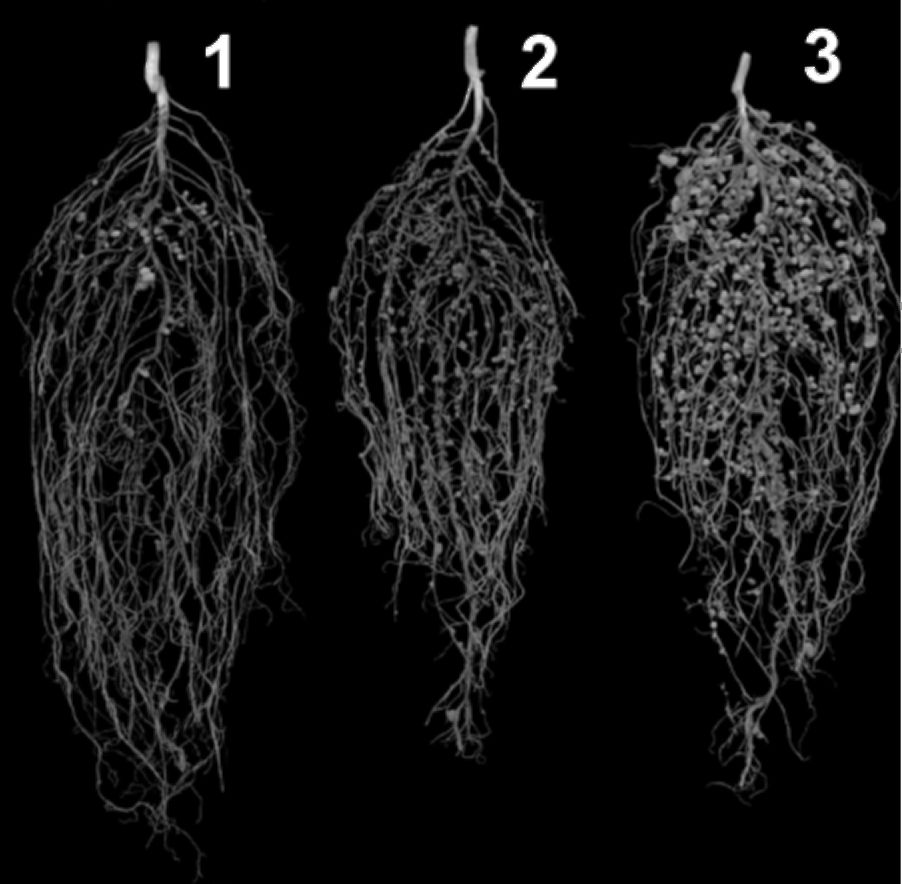
Figure 2. Pea roots: (1) supernodulating; (2) hypernodulating.
17