|
Pisum Genetics |
Volume 24 |
1992 |
Research Reports |
pages 64-74 |
Internode length in Pisum: phenotypic characterisation and genetic identity of the short internode mutant Wt11242.
|
Kusnadi, J., Gregory, M., Murfet, I.C., Ross, J.J., and Bourne, F. |
Department of Plant Science University of Tasmania Hobart, Tas. 7001, Australia |
Fifteen internode length loci have now been established in pea (Pisum sativum L.): Le (10), La and Cry (4, 13), Lm (9), Na (25), Lh, Lk and Ls (14), Lw (7), Lv (18), Lka and Lkb (19), Lkc (21), Lkd (3) and Sln (22). Mutations at eleven of these loci result in short internodes. In some mutant genotypes, e.g. le, na, lh and ls, the short internodes result from a deficiency of endogenous GA1 (6, 12, 17), the principal GA (gibberellin) active in the control of shoot elongation in pea (5). Such mutants respond readily to applied GA1 or GA3. In contrast, other short internode mutants, e.g. lk, lka, lkb, lkc and lkd, do not appear to be deficient in GAs but show a reduced response to applied GA (3, 8, 17, 19, 21). The mutations in these five cases appear to impair functions some distance down the stem elongation transduction pathway from the point of GA reception (2, 8, 15). In addition to short internodes, the mutants lk, lka, lkb, lkc and lkd also share certain other features, e.g. stiff stems and short flower peduncles, which together characterise the phenotypes known as erectoides and semi-erectoides.
In the present paper we report on the characteristics, including GA levels and response to applied GA3, of a short internode pea mutant, Wt11242, and show that the short internodes result from a recessive mutation at the Lka locus.
Materials and Methods
The short internode mutant Wt11242 was selected at the Wiatrowo Plant Breeding Station by Prof Dr W.K. Swiecicki following treatment of the initial line Wt3527 (cv Paloma) with 200 r Nf (fast neutrons) and 0.014% NEU (nitroso ethyl urea). Wt3527 is a dwarf line with genotype le La cryc (I.C. Murfet, unpub.). The following lines held in the Hobart Pisum collection were used in crosses with mutant Wt11242 : L107 (Torsdag, tall = wild type Le La Cry Lm Na Lh Lk Ls Lw Lv Lka Lkb Lkc Lkd Sln), L6 (le crys), L57 (le la cryc lm), L73 (le la), L181 (ls), L206 (lkc), L212 (Cry or cryc lk), L232- (Cry or cryc lv), K29 (lw), K511 (lh) WL1766, (na), WL5835 (lkd), WL5862 (lkb) and WL5865 (lka). The lines carry the wild type dominant allele, except as indicated in parentheses. Further information on these lines may be found in recent publications from this laboratory.
Plants were grown in 14 cm slim line pots in a 1:1 (v/v) mixture of 10 mm dolerite chips and vermiculite topped with 4 cm of sterilised peat and sand potting mix. Nutrient (Hoaglands #1 or "Total Growth Nutrient") was applied once per week. Seed coats were nicked prior to sowing to facilitate imbibition. All plants were grown in our glasshouse and phytotron facility. Most studies (e.g. allelism tests, gibberellin application, determination of GA levels, and anatomical analysis) were conducted using plants grown under an 18 h photoperiod (natural day extended before dawn and after dusk by light from 40 W fluorescent tubes and 100 W incandescent bulbs providing 25 mmol m-2 s-1 at pot top). Data for the Wt11242 x Wt3527 F2 (Table 1 and Fig. 1) were obtained from plants grown under a 15 h photoperiod (9 h daylight + 6 h light from 40 W incandescent globes giving 3 mmol m-2 s-1 at pot top + 9 h dark). Temperature was usually 20-23°C during the 9 h of daylight, and was held at 16°C during the remainder of the 24 h cycle. The plants used for comparison of the growth of mutant Wt11242 and its initial line Wt3527 in darkness (Table 3, Fig. 2) were germinated and grown in a dark chamber held at a constant 21°C.
Nodes were counted from the first scale leaf as node 1. Basal laterals (Table 1) were considered to be those arising from nodes 1-4. Internode 1 lies between nodes 1 and 2.
Anatomical data for the epidermal and outer cortical cells of lines Wt11242 and Wt3527 (Table 2) were obtained using the epidermal strip technique of Arney and Mancinelli (1) and the method of Murfet (11). The means are based on the measurement of 10 epidermal cells and 30 outer cortical cells per plant for each of 8 plants.
Gibberellins were extracted in methanol from the apical portion of the plants above the uppermost expanded leaf as described before (20) and purified for mass spectrometry using high performance liquid chromatography (HPLC) as described by Reid et al. (21). Further harvest details are given in Table 4. The internal standards used were [17-13C, 17-3H2]GA29 (provided by Professor B.O. Phinney, Los Angeles, CA, USA) and [17,17-2H2]GA1, [17,17-2H2]GA8, [17,17-2H2]GA20, and [20-2H1]GA44 (provided by Professor L.N. Mander, Canberra, Australia). The standards were added prior to the commencement of extraction. The quantities added are shown in Table 4. The GAs were quantified by gas chromatography-selected ion monitoring (GC-SIM) following the procedures described previously (8, 21).
The response of lines Wt11242 and Wt3527 to gibberellic acid (GA3) wajs determined by applying 0.1, 1 or 10 ng of GA3 in 10 ml of ethanol to the leaflets of the first foliage leaf (node 3) at the stage when this leaf was completing expansion (day 10). Control plants received 10 ml of ethanol only. Two sets of Wt3527 plants were used: one set was treated on the dry seed with 20 mg of paclobutrazol in 10 ml of ethanol and the other set was not treated in this way. Paclobutrazol inhibits GA biosynthesis and it was used to reduce the internode length of the Wt3527 plants to a value comparable with that of the untreated Wt11242 plants (Fig. 5).
Results
Inheritance and phenotype of mutant Wt11242
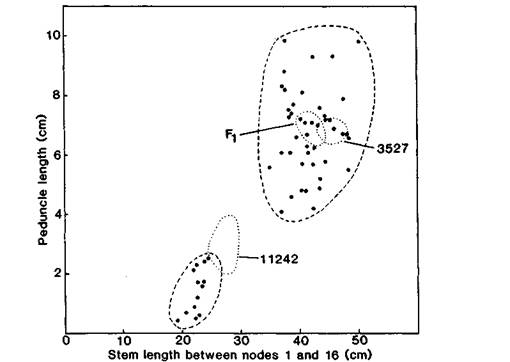
Cross Wt11242 x Wt3527 provided clear evidence of monogenic recessive inheritance of the mutant phenotype. The F1 resembled Wt3527 and the F2 segregated clearly into 44 plants like the initial line and 13 plants like the mutant line (Fig. 1). These F2 numbers are in close agreement with a 3:1 ratio (c2 = 0.15). Mutant plants were characterised by several strongly expressed features including a marked reduction in internode length (reduced 46-48%), final height (reduced 40%), peduncle length of the first flower (reduced 80%) and petiole length of leaf 8 (reduced 48%) (Table 1). Mutant-type segregates could clearly be distinguished on the basis of either stem length between nodes 1 and 16 or peduncle length of the first flower (Fig. 1). The mutant plants also had stiff stems and were wholly devoid of lateral branches in conditions where two thirds of the Wt3527-type segregates produced strong basal laterals (Table 1). These phenotypic traits are characteristic of the erectoides (lk) and semi-erectoides (lka, lkb, Ikc and lkd) mutants (14, 18, 19, 21). Node of flower initiation showed a small (6%) but significant (P <0.01) increase in mutant plants (Table 1).
The reduction in the length of internode 6 of Wt11242 relative to Wt3527 was associated with a significant reduction in the length and number of both epidermal and outer cortical cells (Table 2).
After 13 days growth in the dark, the shoots of Wt11242 were 10% shorter in total length than those of Wt3527 and this difference was significant at the 0.001 level (Table 3). However, there was no significant difference in the length of the epicotyl and internodes 2 and 3, and the difference in stem length was attributable mostly to the fact that 6% fewer leaves were present on the mutant plants but also to the fact that internode 1 was significantly (P <0.01) shorter in Wt11242. Nevertheless, the mutant plants were readily distinguishable from Wt3527 plants on the basis of differences in leaf morphology which are clearly apparent in Fig. 2. While leaf development was suppressed in dark-grown plants of both genotypes, the effect was particularly severe in Wt11242 where the petioles and leaflets remained very small and the leaves were adpressed to the stem (Fig. 2).
Fig. 1. Peduncle length of the first flower plotted against stem length between nodes 1 and 16 for parental lines Wt3527 and Wt11242, and Fl and F2 (●) populations of cross 3527 x 11242. The plots for the parents and Fl fell within the boundaries indicated by the dotted lines. Photoperiod 15 h.
Table 1. Mean ± SE for several traits for Wt3527-type segregates and Wt11242-type segregates in the F2 of a cross between mutant line Wt11242 and its initial line Wt3527, Photoperiod 15 h.
|
Trait |
Wt3527-type F2 plants |
|
Wt11242-type F2 plants |
||||
|
Mean |
SE |
n |
|
Mean |
SE |
n |
|
|
Stem length between nodes 1 and 12 (cm) |
24.75 |
0.39 |
44 |
|
13.29 |
0.34 |
13 |
|
Plant height at maturity (cm) |
79.56 |
1.42 |
44 |
|
47.62 |
1.48 |
13 |
|
Peduncle length of first flower (cm) |
6.64 |
0.25 |
44 |
|
1.43 |
0.21 |
13 |
|
Petiole length of leaf 8 (cm) |
4.04 |
0.06 |
44 |
|
2.10 |
0.10 |
13 |
|
Total length of basal laterals (cm) |
37.32 |
4.86 |
44 |
|
0 |
0 |
13 |
|
Node of flower initiation |
18.47 |
0.15 |
44 |
|
19.54 |
0.29 |
13 |
Table 2. Mean ± SE for cell length and number of cells per internode for epidermal and outer cortical cells for the sixth internode of the mutant line Wt11242 and its initial line Wt3527. n=8
|
Trait |
Initial line Wt3527 |
|
Mutant line Wt11242 |
Significance of difference |
||
|
Mean |
SE |
|
Mean |
SE |
||
|
Length of internode 6 (mm) |
24.50 |
1.30 |
|
9.21 |
0.50 |
P <0.001 |
|
Cell length (mm) |
|
|
|
|
|
|
|
Epidermis |
190.2 |
5.6 |
|
113.5 |
5.7 |
P <0.001 |
|
Outer cortex |
65.3 |
2.3 |
|
49.2 |
1.3 |
P <0.001 |
|
Cell number per internode |
|
|
|
|
|
|
|
Epidermis |
130 |
9 |
|
82 |
6 |
P <0.001 |
|
Outer cortex |
380 |
27 |
|
190 |
14 |
P <0.001 |
Table 3. Mean ± SE for internode length, shoot length, and number of leaves present, for plants of mutant Wt11242 and its initial line Wt3527 grown in the dark for 13 days at 21°C. n=12
|
Trait |
Line Wt3527 |
|
Line Wt11242 |
Significance of difference |
||
|
Mean |
SE |
|
Mean |
SE |
||
|
Length (cm) of |
|
|
|
|
|
|
|
Epicotyl |
5.63 |
0.17 |
|
5.67 |
0.30 |
NS |
|
Internode 1 |
3.95 |
0.13 |
|
2.99 |
0.22 |
P <0.01 |
|
Internode 2 |
9.39 |
0.19 |
|
9.98 |
0.69 |
NS |
|
Internode 3 |
9.04 |
0.22 |
|
9.53 |
0.51 |
NS |
|
Shoot |
38.28 |
0.65 |
|
34.31 |
0.79 |
P<0.001 |
|
Leaves present |
5.14 |
0.13 |
|
4.82 |
0.12 |
NS |

Fig. 2. Mutant line
Wt11242 (right) and its initial line Wt3527 (left) grown in complete darkness
for 13 days at 21°C.
Fig. 3. Mature plants (left to right) of L107 (wild type = tall control), WL5865 (lka, semi-erectoides mutant from L107), the F1 of cross 5865 x 11242, mutant line Wt11242 and its initial line Wt3527 (dwarf le cryc). Photoperiod 18 h.
Allelism tests
Allelism tests were performed against 14 length loci. The results clearly showed that Wt11242 carries a mutation for the Lka gene (Figs 3 and 4). The F1 plants of cross Wt11242 x WL5865 (type line for lka) had slightly longer internodes than either parent but the phenotype retained definite semi-erectoides characteristics and was clearly not wild type (tall). In contrast, F1 plants of crosses between Wt11242 and other erectoides genotypes (lk, lkb, lkc and lkd) showed strong complementation (Fig. 4) and the F1 phenotype was clearly wild type. Likewise the F1s of crosses between Wt11242 and L181 (ls), L232- (lv), K29 (lw) and WL1766 (na) all had a tall phenotype, The F1s of crosses between Wt11242 and lines L6 (le crys), L57 (le la cryc lm) and L73 (le la) all had a normal dwarf (le) phenotype.
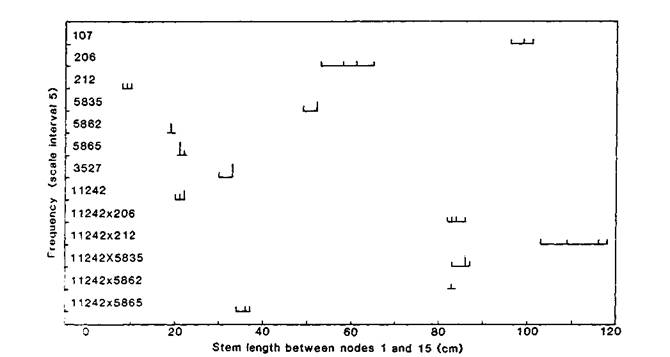
Fig. 4. Distribution of stem length between nodes 1 and 15 for plants of lines L107 (wild type = tall), Wt3527 (dwarf le cryc), L206 (lkc), L212 (Cry or cryc lk), WL5835 (lkd), WL5862 (lkb), WL5865 (lka) and Wt11242 (semi-erectoides mutant from initial line Wt3527), and F1 hybrids from crosses 11242 x 206, 11242 x 212, 11242 x 5835, 11242 x 5862 and 11242 x 5865. Photoperiod 18 h.
Table 4. Level (ng/g fresh weight) of endogenous gibberellins (GAs) detected in the apical buds of 36-day-old plants of mutant Wt11242 and its initial line Wt3527. The material analysed comprised all tissue above the uppermost expanded leaf.
|
Variable |
Initial line Wt3527 |
Mutant line Wt11242 |
|
Plant height at harvest (cm) |
23.06 ± 0.63 |
11.89 ± 0.53 |
|
Leaves expanded at harvest |
9.40 ± 0.16 |
9.30 ± 0.15 |
|
Tissue harvested above node |
9 or 10 |
9 or 10 |
|
Fresh weight of tissue harvested (g) |
16.75 |
15.73 |
|
Number of plants harvested |
32 |
34 |
|
Quantity of internal standard added (ng) |
|
|
|
GA1 |
20 |
20 |
|
GA8 |
25 |
25 |
|
GA44 |
20 |
20 |
|
GA20 |
500 |
500 |
|
GA29 |
500 |
500 |
|
Level of endogenous GA (ng/g FW) |
|
|
|
GA1 |
0.42 |
0.33 |
|
GA8 |
2.10 |
1.65 |
|
GA44 |
0.55 |
0.49 |
|
GA20 |
78.97 |
62.81 |
|
GA29 |
118.60 |
114.33 |
|
GA81 |
1.76 |
1.05 |
Endogenous GA levels
Analysis of endogenous GAs using GC-SIM and labelled internal standards gave no indication of marked differences in GA levels between the mutant Wt11242 and its initial line Wt3527 (Table 4). GA1 is believed to be the major gibberellin active per se in the control of internode length in pea (5). Our results indicate that the level of GA1 and its immediate precursor, GA20, may be about 1.27 times greater in Wt3527 than Wt11242 plants. However, this difference is relatively minor. For example, in the case of the the GA synthesis mutant le, which has a comparable effect to lk11242 on internode length, the level of GA1 was reported to be 10-18 times greater in the wild-type than the mutant plants (24).
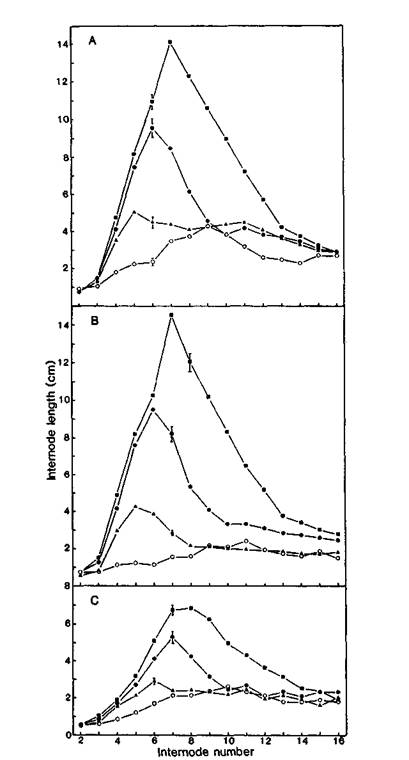
Fig. 5. Internode length plotted against internode number for plants of lines Wt3527 (A, no paclobutrazol; B, treated with 20 mg of paclobutrazol prior to germination) and Wt11242 (C, no paclobutrazol) treated with 0 (○), 0.1 (▲), 1 (●) and 10 (■) mg of GA3 on leaf 3 at day 10. SE bars are fhown in some cases; 60% of SEs were less than 2 mm. Photoperiod 18 h. n = 6-8, mostly 8.
Response to applied GA3
Line 11242 proved much less responsive to applied GA3 than the initial line Wt3527 (Fig. 5). The maximum internode length achieved in Wt11242 following application of 10 mg of GA3 was less than half that achieved in Wt3527 (6.8 versus 14.1 cm). Application of paclobutrazol to the seeds of line Wt3527 reduced the internode length to values comparable with those of the Wt11242 control plants. However, these paclobutrazol-treated Wt3527 plants again responded dramatically to applied GA3, with a maximum internode length of 14.6 cm in the plants given 10 mg of GA3. Thus the lower response of line Wt11242 plants to applied GA3 was not merely a consequence of their short internodes at the time the GA3 was applied.
Discussion
The mutation of gene Lka in the dwarf line Wt3527 to a recessive allele, designated here as lka11242, has resulted in a semi-erectoides phenotype in line Wt11242. In light-grown plants, the mutation caused a significant reduction in internode length, peduncle length, petiole length, the capacity to produce basal branches, and the ability to respond to applied GA3, and a small increase in the node of flower initiation (Table 1, Figs 1, 3 and 5). Endogenous GA levels appeared to be largely unaffected (Table 4). The reduction in internode length was associated with a reduction in both cell length and number of cells per internode for both epidermal and outer cortical cells (Table 2). In dark-grown plants, the difference between genotypes le Lka and le lka11242 was still clearly evident but mostly as a result of a difference in leaf development rather than internode length (Table 3, Fig. 2).
The above effects of allele lka11242 (background le) on internode length, peduncle length, stem anatomy and branching habit are similar to those reported previously for allele lka in a tall (Le) background (18). Thus the deficiency of GA1 caused by the le allele (6) does not prevent expression of this plenotype. The present results also record for the first time effects on petiole length, leaf development in dark-grown plants, and flowering.
A statistically significant, 1-2 node delay in flower initiation in lka11242 plants was observed in several experiments (e.g. Table 1). Line WL5865 (lka) also tended to flower about 2 nodes later than its initial line (L107), e.g. in an 18 h photoperiod WL5865 flowered at node 16.75 ± 0.16 (n=8) and L107 at node 14.67 ± 0.17 (n=9) (P <0.001). Thus there is consistent evidence that these mutations of gene Lka cause an increase of some 6-14% in the node of flower initiation. The erectoides mutant lk possesses elevated ethylene levels (23) and ethylene delays flower initiation in pea (16). Ethylene levels have not been examined in semi-erectoides plants of genotype lka or lka11242 but characteristics consistent with raised ethylene levels were reported for dark-grown plants of WL5865 (lka, 18).
Our results for endogenous GA levels, obtained using labelled standards and GC-SIM, confirm the previous conclusion, based on both similar techniques (8) and bioassays (19), that the effects of mutation of gene Lka are not caused by a reduction in GA levels. It is also clear that the lka11242 allele does not cause an accumulation of 13-hydroxylated C19 GAs, a finding consistent with that reported for lka (8). The levels of the 3b-hydroxylated GAs, GA1 and GA8, reported (8) for mutant line WL5865 (Le lka) and its initial line Torsdag (Le Lka) are some 27 and 17 times greater, respectively, than observed here (Table 4) for Wt11242 (le lka11242) and Wt3527 (le Lka). In contrast, the level of GA20 was 5 times higher in the latter two lines. These differences between the Le lines and the le lines are consistent with the fact that homozygosity for the mutual allele le partially blocks 3b-hydroxylation of GA20 to GA1 (6).
The present results do not permit the relative strengths of mutant alleles lka and lka11242 to be determined with certainty. However, several pieces of evidence support the view that allele lka11242 has a less severe effect than lka. First, in both light-grown and dark-grown plants lka (19 and Fig. 4) reduced internode length relative te its initial line more severely than lka11242 relative to its initial line (Tables 1 and 3, Fig. 4). However, it must be noted that we are comparing lka and Lka on an Le background and lka11242 and Lka on an le background. Second, the F1 plants of cross Wt11242 (le lka11242) x WL5865 (Le lka) had internodes 1.7 times longer than either parent (Fig. 4). This increase above the length of the Le parent could be caused by a general shift in genetic background. However, the fact that the Fl plants of crosses between Wt11242 and other non-allelic, semi-erectoides mutants with a L107 (Torsdag) background, namely L206 (lkc), WL5835 (lkd) and WL5862 (lkb), do not exceed L107 in length gives cause to believe lka11242 is a less severe allele than lka. This view is further supported by the fact that Wt11242 plants consistently flowered 6-7% later than Wt3527 while WL5865 plants flowered 12-14% later than L107 (based on flowering node).
Although alleles lka (19) and lka11242 (Fig. 5) were found to reduce the response to applied GA1 and GA3, respectively, it can be argued that the several effects associated with these alleles are an indication that gene Lka does not act at the point of GA reception but rather at some step further along the transduction pathway to stem elongation (8, 15). The cell wall yield threshold is substantially elevated in lka (WL5865) plants and they may also have a reduced wall yield coefficient (2). However, the primary action of gene Lka remains to be determined.
Acknowledgements. We thank Prof Dr W.K. Swiecicki for providing seed of lines Wt11242 and Wt3527, Professors L. Mander and B. Phinney for providing GA standards, Professor J.B. Reid for comments on the manuscript, Peter Bobbi, Leigh Johnson and Katherine McPherson for technical assistance, and the Australian Research Council for financial support.
Arney S.E. and Mancinelli, P. 1966. New Phytol. 65:161-175.
Behringer F.J., Cosgrove D.J., Reid J.B. and Davies P.J. 1990. Plant Physiol. 94:166-173.
Cramp, R.E. and Reid, J.B. 1992. Plant Growth Regul. (in press).
DeHaan, H. 1927. Genetica 12:321-439.
Ingram, T.J., Reid, J.B. and MacMillan, J. 1986. Planta 168: 414-420.
Ingram, T.J., Reid, J.B., Potts, W.C. and Murfet, I.C. 1983. Physiol. Plant. 59:607-616.
Jolly. C.J., Reid, J.B. and Ross, J.J. 1987. Physiol. Plant. 69:489-498.
Lawrence, N.L., Ross, J.J., Mander, L.N. and Reid, J.B. 1992. J. Plant Growth Regul. 11:35-37.
Lindqvist, K. 1951. Hereditas 37:389-420.
Mendel, G. 1865. Verhand. Naturf. Ver. Brünn 4:3-47.
Murfet, I.C. 1990. Physiol. Plant. 79:497-505.
Potts, W.C. and Reid, J.B. 1983. Physiol. Plant. 57:448-454.
Rasmusson, J. 1927. Hereditas 10:1-150.
Reid, J.B. 1986. Ann. Bot. 57:577-592.
Reid, J.B. and Davies, P.J. 1992. In Karssen, C.M., Van Loon, L.C., Vreugdenhil, D. (eds), Progress in Plant Growth Regulation, Kluwer, Amsterdam, pp 214-225.
Reid, J.B. and Murfet, I.C. 1974. Aust. J. Plant Physiol. 1:591-594.
Reid, J.B. and Potts, W.C. 1986. Physiol. Plant. 66:417-426.
Reid, J.B. and Ross, J.J. 1988. Physiol. Plant. 72:595-604.
Reid, J.B. and Ross, J.J. 1989. Physiol. Plant. 75:81-88.
Reid, J.B., Hasan, O. and Ross, J.J. 1990. J. Plant Physiol. 137:46-52.
Reid, J.B., Ross, J.J. and Hasan, O. 1991. J. Plant Growth Regul. 10:11-16.
Reid, J.B., Ross, J.J., Swain, S.M. 1992. Planta (in press).
Ross, J.J. and Reid, J.B. 1986. Physiol. Plant. 67:673-679.
Ross, J.J., Reid, J.B. and Dungey, H.S. 1992. Planta 186:166-171.
Wellensiek, S.J. 1971. Pisum Newsl. 3:46.