УДК591.174:595.733.4
О. Э. Костерин
НАБЛЮДЕНИЯ ЗА ЛЁТНОЙ АКТИВНОСТЬЮ OPHIOGOMPHUS SPINICORNIS SELYS,
1878 (INSECTA, ODONATA, GOMPHIDAE) В ДОЛИНЕ Р. ШИВЭЭЛИГ-ХЕМ (УБСУ-НУРСКАЯ КОТЛОВИНА,
ЮЖНАЯ ТУВА).
РЕЗЮМЕ.
Приводятся данные о протоколировании трофической лётной активности Ophiogomphus spinicornis Selys, 1878 в низовьях р. Шивээлиг-Хем, Южная Тува, 12 и 15
июля 1990 г., полноценные наблюдения велись 15 июля. В этот день суммарное
время полета составило в бюджете времени 1 %. Распределения вылетов по
длительности и дальности напоминает показательное. Признаков территориализма не
наблюдалось. Наибольшая летная активность наблюдается утром, в интервале
10:00-12:00, тогда же часты смены присады. Во второй половине дня отмечен
полуторочасовой период покоя. Исследуемый вид является наиболее ярким из
известных присадников по системе Ф. Корбета и подстерегателей по системе В. В
Заики.
OBSERVATIONS ON
FLYING ACTIVITY OF OHPIOGOMPHUS SPINICORNIS SELYS, 1878 (INSECTA, ODONATA,
GOMPHIDAE) IN THE SHIVEELIG-KHEM RIVER VALLEY (UBSU-NUR INTERMONTANE HOLLOW,
SOUTHERN TUVA).
O. E. Kosterin
Institute of
Cytology & Genetics SB RAS, Acad. Lavrentyev ave. 10,
Novosibirsk,
630090, Russia. kosterin@bionet.nsc.ru
ABSTRACT. Chronometrical data are presented on
trophical flying activity of Ophiogomphus spinicornis Selys, 1878 at the
Shiveelig-Khem lower reaches, South Tuva, on July 12 and 15, 1990, the complete
data referring to July 15. The total share of flight in the entire time budget
comprised 1 %. Frequency distribution of the flight duration and flight
distance resemble the exponential one. No signs of territoriality were
observed. The largest activity is observed in the morning, at 10:00-12:00 hr,
when perch changes were also frequent. A 1.5 hour long period of rest was
observed in the afternoon. The species studied appears the extreme percher
accoring to Corbet’s system and a watcher according to Zaika’s system.
1. Введение
Являясь
активными воздушными хищниками, стрекозы демонстрируют различные стратегии
поведения, различающиеся в том числе характером и продолжительностью полета. В
1962 г. выдающийся одонатолог Филип Корбет (Corbet, 1962) условно разбил стрекоз на два класса,
«летунов» и «присадников» (fliers
and perchers):
если первые в активной фазе находятся почти в постоянном полете, то вторые
много времени проводят в состоянии покоя на тех или иных присадах, взлетая для
достижения определенной цели (питание, спаривание, агонистические контакты).
Это принципиальное деление принимается и поныне (Corbet, May, 2008). В
нашей стране эти идеи получили развитие в виде системы жизненных форм стрекоз,
предложенной В. В. Заикой (1977, 1979; Заика, Воронова, 1977), который
первоначально выделил три жизненные формы -
преследователи, подстерегатели и собиратели, в дальнейшем дополнив ее
четвертой – патрули (Заика, 1980). Здесь преследователи и подстерегатели более
или менее соответствуют летунам и присадникам Корбета, патрули демонстрируют
как бы промежуточное поведение (таких стрекоз Корбет относит к присадникам, см.
Corbet, May, 2008), а
собиратели, представленные исключительно равнокрылыми стрекозами, используют
длительный активный полет для сбора добычи с субстрата.
Разные
поведенческием модели летной активности должны соответствовать различные
физиологические механизмы терморегуляции (Corbet, May, 2008),
однако этот вопрос недостаточно исследован. Ощущается и недостаток в
фактических количественных данных, касающихся поведения. В Таблице 2 обзора Ф.
Корбета и М. Мэя суммированы данные по летной активности 39 видов, из которых
семейства Aeshnidae, Gomphidae и Corduliidae
представляют лишь по одному виду. Авторы особо оговаривают необходимость
расширения спектра исследованных видов, в особенности за счет гомфид. Данная
работа в какой-то мере восполняет этот пробел, приводя данные индивидуального
протоколирования поведения самцов Ophiogomphus spinicornis Selys, 1878,
проведенные в нижней части долины р. Шивээлиг-Хем. O. spinicornis является малоизвестным китайско-монголским видом, заходящим в Сибирь в
Туве, Прибайкалье и Забайкалье (Костерин, 1999; Kosterin, 2004) (ранее экземпляры с Шивээлиг-Хема ошибочно
приводились В. В. Заикой и О. Э. Костериным (Zaika, Kosterin, 2002)
как Ophiogomphus serpentinus Charp.,
причем сочетающие в себе признаки O.
serpentinus s. str. и O. reductus Calv.);
он включен в первое издание Красной Книги Республики Тыва (Костерин, 2002).
2. Материалы и методы.
Наблюдения
проводились в 1990 г., 12 июля с 9:00 до 12:00 и 15 июля с 9:00 до 18:00, на
обширной поляне в низовьях р. Шивээлиг-Хем (Тес-Хемский кожуун Республики Тыва,
50°45’ с. ш., 94°33’ в. д., около 1300 м над у. м.), по которой были
разбросаны отдельные кусты Caragana arborescens.
Активность стрекозы протоколировалась автором, находившемся в нескольких метрах
от нее, не вызывая беспокойства. Моменты взлета и посадки фиксировались с
помощью электронных наручных часов. Дальность отлета оценивалась на глаз. В
случае потери стрекозы из вида наблюдения продолжались за следующей же
встреченной особью. Наблюдаемые особи являлись самцами, за исключением особи,
наблюдавшаяся с 10:29 12 июля, которая была самкой. Динамика солнечного сияния
отслеживалась при помощи гелиографа. 12 июля солнечное сияние продолжалось в
интервалы 9:14-9:40, 10:02-10:06, 10:13-10:36 и 10:45-10:49, в остальное время
солнце было закрыто плотными облаками, так что в 12 ч наблюдения были
приостановлены. Первая стрекоза была найдена в 9:35, наблюдаемая особь терялась
дважды, наблюдения продолжались вплоть до 12 ч, суммарное время без наблюдаемой
стрекозы составило 11 минут. 15 июля было преимущественно ясно, солнце было
закрыто облаками лишь в интервалы 10:26-10:38, 12:00-12:15, 12:26-12:29,
12:33-14:00, 14:53-14:57, 15:10-15:12, 16:23-16:29, 17:26-17:28, с 17:32-17:36,
17:38-17:57. Первая стрекоза была найдена в 9:43, последняя потеряна в 17:32;
за это время стрекоза терялась 5 раз, суммарное время без наблюдаемой особи
составило 49 минут. Приводится местное летнее время (Гринвич + 8 ч).
3. Результаты
У исследованных особей O. serpentinus отмечены
вылеты двух типов: смены присады и трофические. Первые связаны, скорее всего, с
выбором стрекозой наиболее удобного места и наблюдались в основном утром,
вскоре после выхода солнца, когда стрекозы возвращались с мест ночевки
(предположительно на деревьях) на места дневной охоты. Днем смена присады
наблюдалась редко. В отношении трофических взлетов можно заметить, что в
подавляющем большинстве случаев сидящая стрекоза никак не реагирует на пролетающих
невдалеке насекомых, вполне подходящих на роль добычи. В то же время многие
взлеты в течение дня завершались поимкой добычи. Конспецифический
агонистический контакт наблюдался всего единожды за полтора дня наблюдений,
когда стрекоза взлетела и преследовала подлетевшую особь того же вида. Подобный
пролет мимо других особей наблюдался многократно, но не вызывал вылета
наблюдаемой стрекозы. Наблюдалось также по одному вылету вслед за пролетевшей
мимо Aeshna sp. и ктырем. У стрекоз данного вида совершенно не
наблюдалась такая форма летной активности, как облет индивидуального участка,
равно как и индивидуальных участков как таковых.
Следует заметить, что активная часть вылетов длится
менее секунды, а остальное время, проведенное стрекозой в полете, приоходится
на медленную и «осторожную», в качающемся полете, посадку на присаду, которая
часто не доводится до конца, так как стрекоза перелетает и начинает садиться на
другую присаду.
Однако не следует думать, что большую часть времени
сидящая стрекоза продводит в оцепенении. Она то и дело двигает головой,
сканируя пространство, а также периодически резко поднимает брюшка выше
горизонтальной линии, затем брюшко как бы само постепенно опускается вниз,
после чего стрекоза его снова поднимает. Вращением головы стрекоза реагирует на
всякое движение наблюдателя, находящегося в нескольких метрах. Даже стрекоза,
просидевшая более двух часов после захода солнца за надвинувшийся облачный
фронт, вращала головой и даже протирала глаза передней ногой при попытке наблюдателя
к ней приблизиться.
Присадами для стрекоз чаще всего служили кусты Caragana arborescens. Для 11 случаяев имеется грубая визуальная оценка
высоты стрекозы над землей: 0,5 м – три случая, 1 м – 1 случай, 1,5 м – 4
случая, 2 м – 3 случая. Шесть раз отмечена посадка на стебли травы –
преимущественно сухие соломины, реже – побеги полыни. 15 июля до 11:30
наблюдалась посадка почти исключительно на крупные камни: отмечено 13 таких
посадок, причем это не индивидуальная особенность – за это время сменилось 3 наблюдаемых
стрекозы; позже стрекозы стали садиться на ветки.
Обращает на себя внимание отсутствие взлета
наблюдаемых стрекоз в течение более чем двух часов 12 июля (10:57 - 12:30) и 15
июля (12:25 – 14:32), что с очевидностью объясняется заходом солнца за плотные
тучи. Однако во время периода полного покоя стрекозы в течение ровно полутора
часов, с 15:38 до 17:08 15 июля, небо было чистым, а погода безветренной. Все
это время стрекоза по прежнему двигала головой и периодически поднимала брюшко
вверх). Следует заметить, что это относилось не только к наблюдаемой особи – в
это время не наблюдалось вылетов и у нескольких других особей, находящихся в
поле зрения.
Низкая у данного вида летная активность затрудняет
анализ протоколов, поскольку вылектов и, соответственно, периодов покоя слишком
мало для усреднения по временным интервалам. Кроме того, наблюдаемые особи
иногда терялись, так что в наблюдениях имеются лакуны, общая продолжительность
которых различается в разное время дня. Тем не менее, в Таблице 1 приведены
данные наблюдений 15 июля 1990 г. для интервалов часовой длительности.
Наибольшая частота вылетов наблюдалась в интервале с 10:00 до 11:00, в течении
которого было совершено 14 вылетов, 12 из которых сопровождались сменой
присады. В следующий интервал, с 11:00 до 12:00, вылетов было зафиксировано
немногим меньше, 12, из них 10 сопровожалось сменой присады. В этот же интервал
наблюдалась наибольшая средняя продолжительность вылетов, 13,7 с., при разбросе
наблюдавшихся значений этого параметра от 1 до 52 с. В остальные часовые
интервалы, в которые летная активность имела место, средняя продолжительность
полета варьировала около 4 с, хотя объемы выборок крайне невелики чтобы делать
какие-либо сравнения между ними. Наибольшая средняя дальность вылета 3,8 м (при
разбросе 0,3-10 м) зафиксирована утром, в интервале 10:00-11:00, затем этот
параметр упал до 1-2,5 м.
12 июля в единственный часовой интервал 10:00-11:00,
когда имели место солнечное сияние (общей продолжительностью 29 мин) и
активность стрекоз, было зафиксировано 8 вылетов (из них 6 со сменой присады),
со средней длительностью 15 с и средней дальностью 2,2 м.
Доля полета в общем бюджете времени рассчитана для
полного дня наблюдений 15 июля, отличавшегося преимуществено хорошей погодой. В
сумме время непосредственного наблюдения за отдельной стрекозой (за вычетом
времени поиска следующей особи после потери предыдущей из вида) составило 414
минут (помимо периода, охваченного в Табл. 1, учтены наблюдения в период с 9:47
до 10:00, в течение которого вылетов не наблюдалось). Из них в полете стрекоза
находилась в сумме 249 секунд, что составляет 1,0 % времени. Если отнести
суммарное время полета к сумарному времени наблюдений при прямом солнечном
сиянии, составляющему 260 минут, то получим цифру в 1,6 %. Полеты,
завершившиеся потерей особи из вида или появлением новой наблюдаемой особи,
учтены лишь в 4 из 12 случаев, поэтому рассчитанная средняя длительность полета
несколько занижена, но никак не более чем вдвое.
На Рис. 1 приведена гистограмма распределения
вылетов по их продолжительности, а на Рис. 2 – по дальности отлета. Среднее
время полета получается 4,87 с, его среднеквадратическое отклонение – 3,29 с (n=46). Отметим, что эти параметры слабо различаются,
что может указывать, что само распределение близко к показательному, которое
оно напоминает также и по форме. При подсчете этих показателей были отброшены
два явно выбивающихся значения: 33 с и 52 с. Во время первого из этих вылетов
стрекоза дважды примеривалась сесть: на землю и на соседний куст караганы, но
затем вернулась на тот куст, на котором она сидела ранее. Во время второго
вылета стрекоза пыталась сесть в район, где уже сидела другая особь, и была
прогнана ею (это был упоминавшийся выше единственный агонистический контакт).
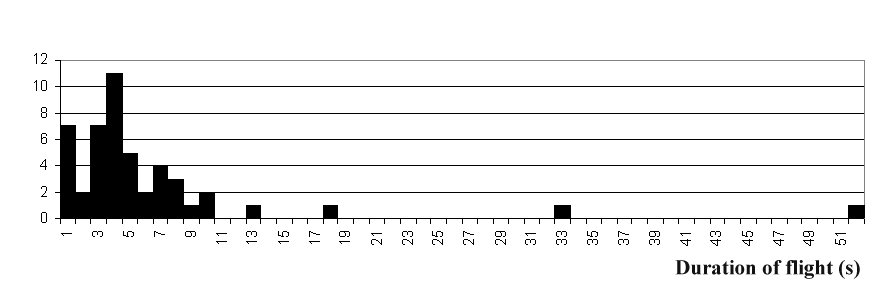
Рис. 1. Гистограмма распределения числа полетов по их продолжительности
(интервалы по 1 с).
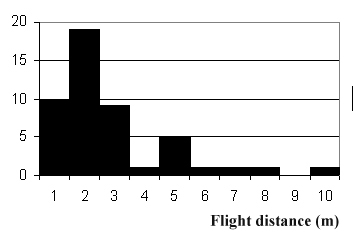
Рис. 2. Гистограмма распределения числа полетов по их
дальности (интервалы по 1 м).
Средняя дальность отлета получилась 2,4 м,
среднестатистическое отклонения – около 2 м. Снова отметим близость этих
показателей распределения. К этим данным следует относиться с осторожностью,
так как они основаны на визуальной оценке дальности. Кроме того, здесь имеется
систематическая ошибка, состоящая в том, что во время наиболее дальних отлетов
стрекоза терялась из поля зрения, так что они не попадали в протокол (в
какой-то мере это касается также данных по времени полета).
4. Обсуждение
Прежде
всего отметим, что мы фактически фиксировали трофическую активность стрекозы,
поскольку данный вид стрекоз развивается в реке Тес-Хем, но не в слишком
быстром и холодном Шивээлиг-Хеме, так что наблюдаемые особи находились за
пределами своих репродуктивных сайтов. В то же время наблюдения, суммированные
в обзоре Корбета и Мэя (Corbet, May, 2008), делались преимущественно в репродуктивных
сайтах, где смешивается репродуктивная и трофическая активность. Данное
обстоятельство, с одной стороны, является преимуществом, так как нами
исследовалась только одна форма активности в чистом виде; с другой стороны, оно
затрудняет сопоставление с данными, полученными другими авторами. Сугубо
трофический характер наблюдавшейся активности подтверждается крайней редкостью
агонистических контактов (всего три случая, если считать за таковые вылет вслед
за коромыслом и ктырем).
Полученные
данные недостаточны для надежных количественных оценок лётной активности
данного вида стрекозы, так как представляют один полный день наблюдений, что не
позволяет провести усреднений по многим индивидуумам в один и тот же интервал
времени и тем самым отбросить возможное влияние индивидуальных особенностей
наблюдаемых стрекоз. Однако они достаточны для иллюстрации того очевидного
факта, что O. spinicornis, подобно всем сибирским гомфидам, является ярчайшим
представителем присадников (подстерегателей). (В более южных широтах имеются
гомфиды с гораздо большей летной активностью, например, Sieboldius.) По нашим данным, в бюджете времени этого вида полет
составляет почти точно 1 %, если учесть только суммарное время прямого
солнечного сияния, то получим 1,6 %. Эта цифра несколько занижена, так как не
учтены взлеты, окончившиеся потерей стрекозы из виду наблюдателем. Заметим, что
имеющиеся в литературе количественные данные по единственному виду гомфид,
американскому Argiogomphus villosipes (Selys)
демонстрируют долю полета в бюджете времени в 2% (McMillan, 2006; цит. по Corbet, May, 2008).
Таким образом, среди всех исследованных видов стрекоз на данный момент O. spinicornis обладает
наименьшей летной активностью. Такая низкая лётная активность сочетается с ее
довольно высокой трофической эффективностью. Для данного вида совершенно
нехарактерен облет индивидуального участка, который в той или иной степени
демонстрируют многие присадники из семейства Libellulidae и который является основной формой активности у
летунов. Более того, у O. spinicornis мы вообще не выявили каких-либо признаков
существования территориализма.
Корбет
и Мэй (2008) поднимают проблему что считать активностью стрекозы, находящейся
на присаде, и предлагают сохранение осторожности в качестве критерия того, что
стрекоза «активна». Наши наблюдения, касающиеся движений сидящей стрекозы,
свидетельствуют о том, что наблюдаемые стрекозы были без сомнения активными.
Именно
длительные периоды покоя стрекозы представляют наибольший интерес. Тогда как
двухчасовые периоды покоя 12 июля объясняются заходом солнца за плотные тучи,
что совершенно блокирует летную активность этого вида, трудно найти объяснение
полуторачасовому периоду покоя при благоприятных погодных условиях,
наблюдавшемуся 15 июля (который распространялся и на соседних особей). Все это
время стрекоза по прежнему двигала головой и периодически поднимала брюшко
вверх). Следует заметить, что отсутствие лётной активности относилось не только
к непосредственно наблюдаемой особи – в этот период времени не наблюдалось
вылетов и у нескольких других особей, находящихся в поле зрения. Во-первых, не
исключено, что стрекозы к этому времени насытились и прекратили трофические
вылеты, вновь приобретя летную активность перед тем, как долину накрыла тень от
горы, то есть перед отлетом на ночевку. Во-вторых, это было наиболее жаркое
время дня, и покой стрекоз мог быть мерой против перенагрева.
Распределение
полетов по их длительности напоминает показательное как по форме, так и по
близости значений среднего и среднеквадратического отклонения. Время полета
было бы в точности распределено показательно, если бы стрекоза в любой момент
полета могла бы его с одной и той же вероятностью прекратить, вне зависимости
от того, сколько времени она находилась в полете до этого. Для такого допущения
у нас, казалось бы, нет особых оснований. У полета есть определенная цель
(поимка добычи), по достижении которой его продолжение лишено особого
биологического смысла. По-видимому, должен существовать также и оптимальный
радиус расстояний до добычи, которая стимулирует вылет, и, соответственно,
оптимальное время, необходимое для ее поимки. Однако выше было отмечено, что
собственно поимка добычи занимает мало времени, тогда как основное время полета
тратится на посадку. Последнее может зависит от возбужденности стрекозы. Но мы
не можем исключить также существование некоего нервного триггера, переключающее
у стрекозы режимы полета и покоя, который по достижении цели полета может
срабатывать более или менее случайно. Это могло бы объяснить показательный
характер наблюдаемого распределения.
Наблюдавшиеся
стрекозы демонстрировали элементы пассивной терморегуляции, характерные именно
для присадников (Corbet, May, 2008). Сюда относится тот факт, что до 11:30 15 июля
они садились исключительно на крупные камни, по всей видимости уже нагретые
солнцем, после чего стали использовать в качестве присад ветки. Длительный
период покоя во второй половине дня, то есть в наиболее жаркое время дня, также
можно интерпретировать как способ избегать перегрева. Периодические поднятия
брюшка, возможно, также связаны с терморегуляцией, однако поза обелиска как
таковая не отмечена. Послеполуденный покой, как отмечалось, также мог быть
элементом пассивной терморегуляции.
5. Благодарность.
Автор
выражает благодарность В. В. Заике за возможность принять участие в комплексной
биосферной экспедиции в Южной Туве в 1990 г.
6. Литература.
Заика В. В. 1977. Поведение стрекоз Северной Кулунды в
репродуктивный период. // Этологические проблемы экологии насекомых Сибири.
Новосибирск: изд-во НГУ, с. 108-125.
Заика В. В. 1979. Население стрекоз наземной части
биогеоценозов Северной Кулунды. // Вопросы экологии. Новосибирск: изд-во НГУ,
с. 87-111.
Заика В. В. 1980. Экология и поведение стрекоз (Odonata) острова Кунашир. // Вопросы экологии.
Новосибирск: изд-во НГУ, с. 73-88.
Заика В. В., Воронова И. А. 1977. Поведение стрекоз в озерной степи
Западной Сибри. // Этологические проблемы экологии насекомых Сибири.
Новосибирск: изд-во НГУ, с. 82-105.
Костерин О.Э. 1999. Фауна стрекоз (Odonata) Даурского заповедника и его
окрестнотей. // Насекомые Даурии и сопредельных территорий (Сборник научных
трудов). Выпуск II - Новосибирск - С. 5-40.
Костерин О. Э. 2002. Дедка шипорогий Ophiogomphus spinicornis Selys, 1878. // Красная книга республики Тыва.
Животные. Изд.-во СО РАН, филиал ГЕО, Новосибирск. - с. 14.
Corbet, P. S. 1962.
A Biology of Dragonflies. Whiterby, London.
Corbet, P. S., M.
L. May. 2008. Fliers and perchers among Odonata: dichotomy or multidimensional
continuum? A provisional reappraisal. // International Journal of Odonatology
11 (2): 155-171.
Kosterin, O.E., 2004.
Odonata of the Daurskiy State Nature Reserve area, Transbaikalia,
Russia. // Odonatologica 33: 41-71.
McMillan, V. E.
2006. Preliminary observations of reproductive behaviour in Argiogomphus
villosipes (Selys) (Anisoptera: Gomphidae). // Bulletin of American
Odonatology 10: 19-22.
Zaika, V.V.,
O.E.Kosterin. 1992. Some interesting observations of dragonflies (Odonata) in
South Tuva. // Acta Hydroentomologica Latvica 2: 81-84.
Костерин Олег Энгельсович, Институт цитологии и
генетики СО РАН, пр. акад. Лаврентьева 10, Новосибирск 630090. kosterin@bionet.nsc.ru
Таблица
1. Лётная активность Ophiogomphus spinicornis в пределах часовых
интервалов 15 июля 1990.
|
Интервал времени |
Кол-во последо- вательно наблю-давшихся особей |
Полное время наблюдения (мин) (в скобках – кол-во набл. особей) |
В том числе при открытом солнце (мин) |
Число вылетов |
Число вылетов со сменой присады |
Число вылетов с поимкой добычи |
Среднее время полета (c) |
Средняя даль-ность полета (м) |
Число посадок на камни |
|
10:00-11:00 |
2 |
53 |
48 |
14 |
12 |
1 |
3,9 (n=13)1 |
3,8 (n=13) |
10 |
|
11:00-12:00 |
4 |
33 |
33 |
12 |
10 |
1 |
13,7 (n=7) |
1,8 (n=6) |
3 |
|
12:00-13:00 |
1 |
43 |
13 |
5 |
2 |
2 |
5,2 (n=5) |
0,9 (n=4) |
0 |
|
13:00-14:00 |
1 |
60 |
0 |
0 |
0 |
0 |
- |
- |
0 |
|
14:00-15:00 |
1 |
60 |
58 |
6 |
0 |
0 |
4,0 (n=6) |
2,3 (n=5) |
0 |
|
15:00-16:00 |
1 |
60 |
2,5 |
8 |
5 |
1 |
4,1 (n=8) |
1,6 (n=8) |
0 |
|
16:00-17:00 |
1 |
60 |
55 |
0 |
0 |
0 |
- |
2,6 (n=4) |
0 |
|
17:00-18:00 |
1 |
32 |
30 |
5 |
3 |
|
4,5 (n=4) |
|
0 |
1 n – число вылетов с измеренной длительностью или дальностью (сюда могут входить и вылеты, сопряженные с потерей наблюдаемой особи, если она садилась в неудобном для наблюдения месте или взлет новой наблюдаемой особи был зафиксирован).
ПОДПИСИ К РИСУНКАМ