plates. The plates were incubated for eight days at 23°C under continuous florescent cool white 34-watt lights. Following incubation, three plugs (4 mm in diameter) containing mycelia were transferred from the agar plates into 120 ml flasks containing Kerr's Medium (15). The flasks were placed on a shaker for seven days at 23°C under continuous florescent cool white light. Cultures of each isolate were then strained through one layer of cheesecloth and centrifuged at 2500 rpm for 5 minutes. The supernatant was poured off, and the spores were re-suspended using sterilized distilled water. The conidia concentrations for each isolate were determined using a hemacytometer, and the suspensions were adjusted to 1 x 106 conidia per/ml of water. Prior to inoculation, equal volumes of the isolates were combined into a single suspension and the seeds were immediately inoculated.
Fifty seeds of each pea genotype were placed in each of two 100 ml beakers to which 60 ml of a conidial suspension of Fsp were added to one beaker and 60 ml of sterilized distilled water were added to the other. The seed were soaked for 17 h at 25°C. The seed of each genotype soaked in sterilized distilled water were used as non-inoculated controls. Following the soaking period, ten wet seeds of each inoculated and non-inoculated genotype were planted in separate plastic trays (52.5 by 26.25 by 6.25 cm) filled with propagation grade-course perlite (Supreme Perlite Company, Portland, OR). Each tray contained three inoculated or three non-inoculated pea genotypes, each planted in the trays lengthwise in single rows with 10 plants per row. There were three replications of each genotype per test that were inoculated and non-inoculated, and each test was repeated one or two times for a maximum of three separate screening tests per genotype. Due to greenhouse space constraints, the forty-four accessions screened for Fsp resistance were divided into four tests labeled 1 through 4, with eleven different PI accessions screened per test. Multiple screenings of the same genotypes conducted at different times were identified as Test 1a, Test 1b and Test 1c. In addition to the eleven accessions screened per test, each test also contained two Fsp-susceptible pea cultivars (Dark Skin Perfection and Bolero) and two Fsp-moderately-resistant PI accessions (PI257593 and PI166159) as controls, except in Test 1a and 1b, where only Dark Skin Perfection was used as a control.
The plants were grown in a greenhouse under natural sunlight, and supplemental lighting with 1000-watt metal halide lights used as needed to maintain a photoperiod of approximately 14 hours. The greenhouse temperatures for each test ranged from 15 to 28°C. The plants were watered uniformly as needed with approximately 500 ml of water per tray and fertilized uniformly with 500 ml of Miracle-Gro (24-8-16, N-P-K) (Marysville, OH) at a concentration of 4.93 ml Miracle-Gro per liter of water at 9, 12, 15, 18 and 21 days after planting.
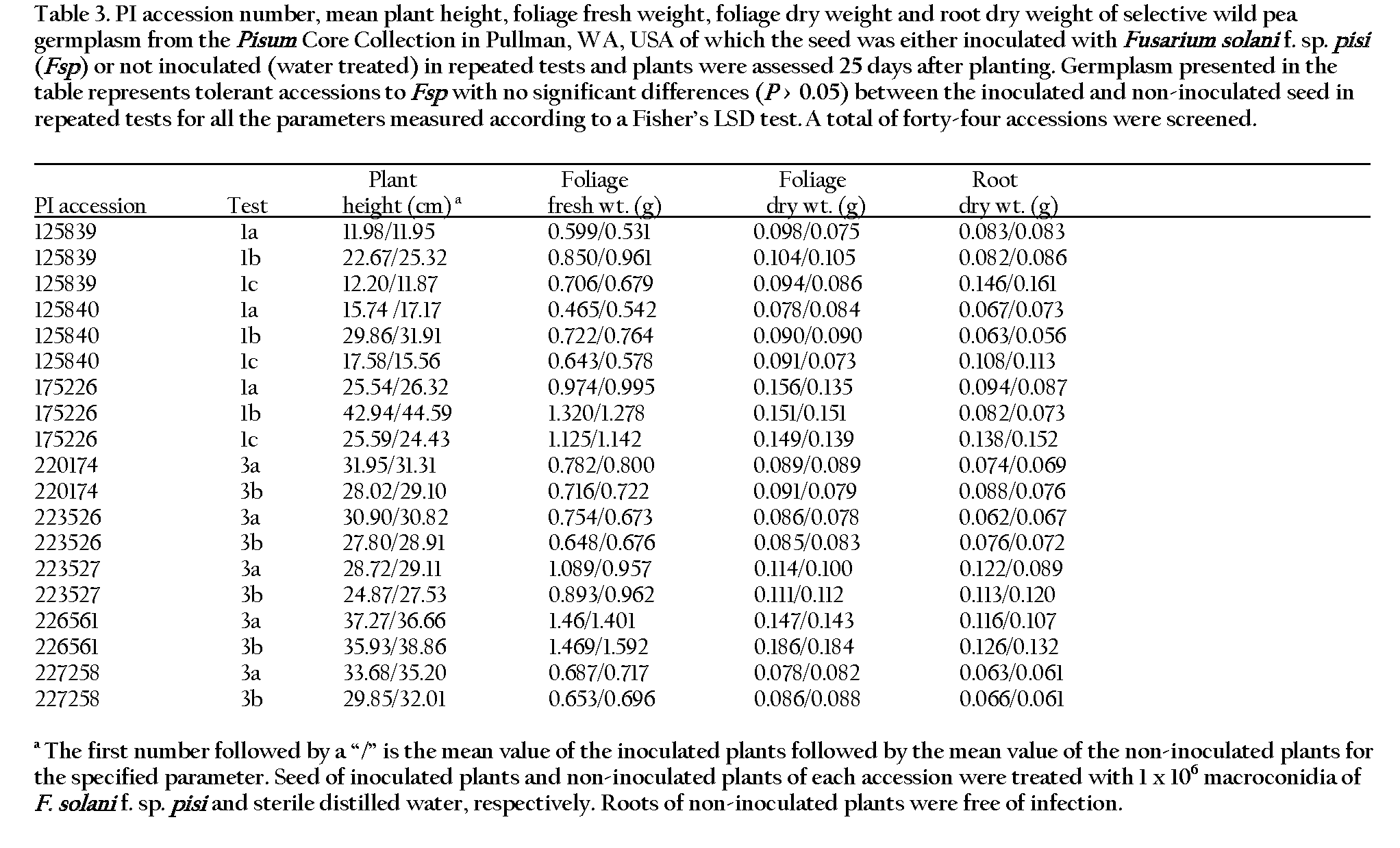
The pea plants were harvested 25 days after planting, which allowed sufficient time for root rot symptoms to develop, and disease resistance was characterized based on the following comparisons between inoculated and non-inoculated plants of each accession: 1) percent germination of seed, 2) root disease