
A floral homeiotic mutation affecting B function of florogenesis control in the pea
| Kosterin, O.E | Institute of Cytology and Genetics |
In 1997 we undertook an extensive experiment on EMS mutagenesis with our pea hybrid Tau2 being maintained as a heterozygote for the Hammarlund translocation, which is described in this issue by Gorel et al. (3). In another communication in this issue (5) we have described a lethal induced in linkage group I which was identified in one of the M3 families during the course of this experiment. Four plants of the same M3 family, which lacked the cri phenotype, appeared to have another recessive mutation with a very drastic effect on flowers. It was not linked to the breakpoint but was inherited by some plants in a heterozygous state; it appeared as a homozygote in about 1/4 of the plants in two subsequent M4 and M5 generations.
The mutation obtained should be regarded as homeiotic. At first glance the flowers of the mutant plants resemble clusters of pistils surrounded by a double whorl of sepals (Fig.1).
Fig. 1. Typical floral phenotype of the multi-pistillate mutation described in text.
The mutant flower lacks normal petals and stamens. The second whorl of five green sepals substitutes for the whorl of petals except that the inner sepals lie directly in front of the outer sepals rather than alternating with them. The stamens are replaced by highly variable structures which share the characters of sepals, pistils and, sometimes, stamens. Each flower usually has several sepal-like structures with one of the margins rolled to resemble a carpel with an apical, often slightly reduced stigma (occasionally these structures resemble normal sepals or have one margin only slightly swollen). Each flower also has a few stamen filaments without anthers (very rarely normal stamens). Several cases were observed of pistillate structures possessing two anthers and fertile pollen at the base of the style (Fig. 2). The number of structures in the stamen whorl is difficult to determine as some of them seem to be fused at their base or to bifurcate distally, but as a rule there are 8 to10 of them. Lastly, each flower bears a normal pistil in the proper position. The flowers do not fade for a long time, the pistil and pistil-like structures grow to 1-3 cm in length and become swollen (Fig. 3). The mutant plants produce no seeds and thus grow much higher and more luxuriant than their normal counterparts.
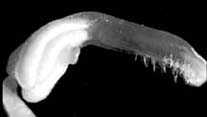


Fig. 2 (left). Enlargement of a pistillate structure from the mutant
line showing two anthers at base of pistil.
Fig.3a, b (center, right). Flowers from the mutant line showing the relatively large
sterile pistils.
We undertook 60 attempts to pollinate the mutant flowers with pollen from the tester line WL1238. From these crosses we obtained nine pods from which 13 seeds were collected. In each flower only one pistil produced seeds, presumably the normal one. The nine F
11 plants were selfed in the greenhouse, and the F2 population (144 individuals) was planted in the field. Joint segregation analysis was performed with the mutant phenotype and the segregating genes le, d, s, wb, k, tl, r, u, m, fl. The mutation exhibited no linkage with any of these markers.A similar mutation has been described by Muehlbauer and Konzak (6), in which the flowers had 8-10 pistils as “the staminate floral parts had differentiated into pistillate parts.” Although the photograph much resembles our phenotype, it was stated that “the number of floral petals remained unchanged,” while in our case they completely turned into sepals. Allelism tests have not been performed between our mutant and that of Muehlbauer and Konzak (6). We do not believe our mutant is an allele of cc (calyx carpellaris), another mutation seriously affecting the flower morphology, becaue cc was shown to be linked to R (7), while ours does not display linkage with this locus. In addition, the nature of abnormalities is quite different in these two mutations.
The genetic control of the floral organ development by a system of homeiotic genes has been well studied in Arabidopsis and Anthyrinum and was shown to be very similar in these distant taxa (for a review see ref. 2). In short, there are three general functions, A, B and C, expressed by corresponding homeiotic genes in a normal flower meristem in the four subsequent whorls providing, respectively, 1) sepals, 2) petals, 3) stamens, and 4) pistils. Function A is expressed in whorls 1 and 2, function C in whorls 3 and 4. These two functions replace each other. For instance, if a mutant lacks function A then function C is expressed in all the four whorls, and vice versa. The function B is expressed in whorls 2 and 3. Function A alone determines development of the primordia into sepals, a combination AB produces petals, the combination CB leads to stamens, and function C alone leads to pistils. If some functions are eliminated by mutations, the proper floral organs in the four whorls are replaced by organs of other types, in accordance with presence of the remaining functions.
There are a number of mutations in pea that have significant effects on the flower structure (1), but so far only one fits the Arabidopsis model. This mutant is cc, described by Gottschalk (4), which causes a reduction of the corolla and turns sepals into pistil-like structures. This mutation appears to eliminate function A. Hence, in the four whorls the following functions are expressed: C, CB, CB, C, resulting in whorl 1 developing the same organs as whorl 4 (pistils) and whorl 2 developing the same organs as whorl 3 (stamens). This pea mutation may correspond to the mutations ovu in Antirrhinum and ap2 in Arabidopsis (2).
The mutation described in the present communication appears to switch off function B, making whorl 2 identical to whorl 1 (function A, sepals) and whorl 3 the same as whorl 4 (function C, pistils). The change of identity of whorl 3 is not complete in this mutant, as some structures resemble stamen filaments or remain normal stamens in some cases. Perhaps function B is not completely eliminated in this whorl. In any case, our mutation may correspond to the mutations def, glo or sep in Antirrhinum and pi or ap3 in Arabidopsis (2). The identification of this mutation as well as calyx carpellaris in pea suggests that the pattern of genetic control of floral morphogenesis in the Fabaceae is principally the same as it is in quite distant families such as Scrophulariaceae and Brassicaceae.
Acknowledgement: This work was partly supported by the Russian State Program, “Russian Fund for Fundamental Research.”
1. Blixt, S. 1972. Agri Hortique Genetica 30:1-293.
2. Coen, E.S. 1991. Annu. Rev. Plant. Physiol. Plant Mol. Biol. 42:241-279.
3. Gorel, F.L., Kosterin, O.E., Berdnikov V.A. 1999. Pisum Genetics 31:5-8.
4. Gottschalk, W. 1964. Bot. Stud. 14:1-359.
5. Kosterin, O.E., Berdnikov, V.A. and Gorel F.L. 1999. Pisum Genetics 31:11-12.
6. Muehlbauer, F.J., Konzak, C.F. 1973. PNL 5:33.
7. Myers, J.R., Gritton, E.T. and Struckmeyer, B.E. 1984. Crop Science 24:1063-1069.