Inheritance and allelism tests on six further branching mutants in pea
Symons, G.M. and Department of Plant Science
Murfet, I.C. University
of Tasmania
Hobart,
Tas. 7001, Australia.
A number of ramosus mutants with increased branching and clear Mendelian inheritance have now been identified in the pea. These include ram (11,12), rms1 (7), rms2 (2), rms3 (2), rms4 (2) and rms5 (1,2). In addition , two genes Fr and Fru , with a quantitative effect on branching have been proposed on the basis of the arbitrary partitioning of continuous unimodal F2 distributions for number of stem branches (6,10). Internode length and flowering genes also influence the branching habit in the pea (2,8,9,13,14). Mutations, such as le (dwarf), which diminish internode length by blocking gibberellin synthesis increase the tendency to produce basal laterals (secondary stems) while mutations which shorten internode length by blocking steps after gibberellin reception reduce the occurrence of lateral branching (2,9,14). Among the flowering genes, those which control the ability to respond to photoperiod have a major effect (8,9,14). In wild-type (WT) lines, which are photoperiodically responsive, the propensity to produce basal laterals increases markedly under short day conditions. Mutations in any of the three complementary photoperiod response genes Sn, Dne and Ppd lead not only to a day neutral flowering habit but also to a diminished capacity to produce basal laterals under any conditions.
Clearly expressed mutations like those of the rms series offer an excellent opportunity to explore the physiological and biochemical control of apical dominance. Results from reciprocal grafts between mutant and WT isolines indicate that the Rms2 gene may control production of a graft-transmissible hormone-like substance which suppresses outgrowth of axillary buds (4). Under an 18-h photoperiod, branching in an rms2 (K524) shoot was completely suppressed by grafting to a WT rootstock. Likewise, the Rms3 gene also appears to influence the production or release of a graft-transmissible substance as the onset of aerial lateral production (branching from the upper nodes) was significantly delayed in mutant rms3 (K564) scions grafted to WT rootstocks (5). In contrast, the phenotype of an rms4 (K164) shoot was unaffected by grafting to a WT rootstock, i.e. the Rms4 gene appears to act entirely in the shoot (5). Nevertheless, recent data obtained using this rms4 mutant show that the genotype of the pea shoot exerts a profound influence over the export of cytokinins in the root xylem sap (3). In particular, the level of the important cytokinin zeatin riboside was found to be some 40-fold greater in the root xylem sap from WT plants than from rms4 mutant plants. Moreover, results from reciprocal grafts between WT and rms4 plants showed that rms4 roots exported normal levels of cytokinins when grafted to a WT shoot but export of zeatin riboside from a WT rootstock was diminished over 40-fold by grafting to an rms4 shoot compared with the level for WT/WT self-grafts, i.e. the level of cytokinins in the root xylem sap was determined by the genotype of the shoot.
While the effect of the rms2 and rms3 mutations on cytokinin levels has not been reported, it is known that both mutations significantly influence auxin levels. Indole-3-acetic acid levels were found to be up to 5-fold higher in rms2 (K524) plants than in WT controls while the levels in rms3 (K564) plants were up to 2-fold higher than in WT controls (4,5). In contrast, indole-3-acetic levels were not significantly altered in the rms4 (K164) mutant (5).
We report here on the inheritance and allelism of six additional ramosus mutants . All showed clear expression and single gene recessive inheritance. Four proved to be allelic with rms1 and two with rms4.
Material and Methods
The six new ramosus mutants M2/8, M2/355, M2/549, M2/716, M3/129 and XV/23 were induced and selected at the Institute of Plant Genetics, Sofia, Bulgaria. Further details are given in Table 3. Inheritance was tested by crossing each mutant with the initial line. Allelism was tested by crossing among the six mutants and by crossing each mutant with the type lines (see 2) for rms1 (WL5237), rms2 (K524), rms3 (K487), rms4(K164) and rms5 (Wt15244). In total 45 crosses were made to test allelism. All lines used have a WT flowering habit as defined by Murfet and Reid (13). Lines WL5237, K524, K487, K164 are tall (Le). Lines Wt15244, Borek, Auralia, XV/23, M2/8, M2/355, M2/549, M2/716, L9-76 and M3/129 are dwarf (le).
The plants were grown in 14-cm slimline pots in a 1:1 (v:v) mixture of vermiculite and 10-mm dolerite chips topped with 3 to 4 cm of pasteurised peat/sand potting mixture. Seeds were sown at a rate of one (allelism tests) or two (inheritance tests) per pot. Nutrient solution was supplied once a week in the form of Total Growth Nutrient (R&D Aquaponics, Sydney) or Aquasol (Aquasol Hortico Ltd, Melbourne). Plants were grown in the glasshouse under an 18-h photoperiod consisting of the natural daylight extended morning and evening by light from fluorescent tubes (Wotan 40W cool white) and incandescent globes (Sylvania 100W) providing an intensity of about 25 µmol m-2 s-1 at pot top.
The degree of lateral branching for each plant was measured by the ratio TLL:TL where TL is the total length of the main shoot and TLL the combined total length of all lateral shoots. This index permits valid comparison between tall and dwarf plants (see 2).
Table 1. Results for the F1 and F2 of crosses between the six new branching
mutants and their initial lines. Phenotype : WT, wild type ; M, mutant.
| Cross | F1 | F2 segregation
WT M |
Chi-square
testing 3:1 |
Probability |
|
M2/8 x Borek |
WT | 42 14 | 0.00 | P=1 |
| M2/355 x Borek | WT | 48 12 | 0.80 | P>0.3 |
| M2/549 x Borek | WT | 38 14 | 0.10 | P>0.7 |
| M2/716 x Borek | WT | 44 15 | 0.01 | P>0.9 |
| M3/129 x L9-76 | WT | 35 17 | 1.64 | P=0.2 |
| XV/23 x Auralia | WT | 44 13 | 0.15 | P=0.7 |
Results
The six new mutants proved readily amenable to Mendelian analysis. Under the 18-h photoperiod, each of the mutants branched extensively at basal and aerial nodes. Lateral outgrowth generally did not occur at nodes 4 to 7. The mutant plants therefore exhibited the G or gap branching pattern described by Arumingtyas et al. (2). The initial lines showed little or no branching under the 18-h photoperiod. Crosses between the mutants and their initial lines produced F1 plants with a WT phenotype and in the F2 generation segregation into the WT and mutant classes was visually distinct with the observed numbers in good accordance with a 3:1 ratio in each cross (Table 1). The ramosus phenotype of mutant plants was usually clearly evident by age 4 to 5 weeks, well before the appearance of open flowers. Thus each mutant showed clear expression and single gene recessive inheritance.
The results of allelism tests (Table 2 and Fig.1) showed that the four mutants M2/8, M2/355, M2/549 and M3/129 are allelic with each other and with rms1 (WL5237). The remaining two mutants, XV/23 and M2/176, proved to be allelic with each other and rms4 (K164).
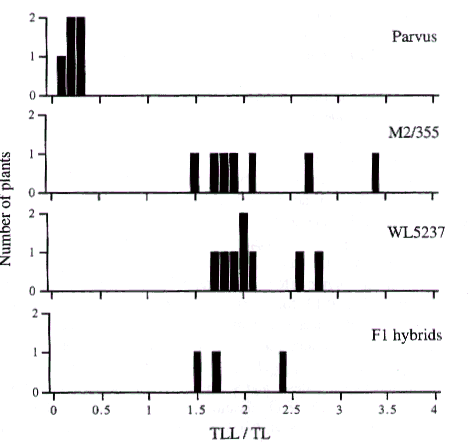
In all crosses testing allelism, the F1 plants were either clearly mutant or clearly WT. Nevertheless, measurements were made on all plants and typical data from these crosses for two allelic mutants are exemplified in Fig.1 by the results for the cross M2/355 ґ WL5237 (rms1).
Fig.1. Distribution of the index TLL:TL (ratio of total lateral length to total length of the main shoot) for 80-day-old plants of Parvus (the WT initial line for WL5237), mutant lines M2/355 and WL5237 (rms1), and the F1 of the cross M2/355 ґ WL5237. Borek, the WT initial line for M2/355, did not branch. Photoperiod 18 h.
Table 2. Results of crosses among the six new ramosus mutants and between each of the new mutants and the type lines for rms1 (WL5237), rms2 (K524), rms3 (K487), rms4 (K164) and rms5 (Wt15244). A, allelic, F1 had mutant phenotype ; NA, not allelic, F1 had WT phenotype.
| WL5237 | K524 | K487 | K164 | Wt15244 | M2/8 | M2/355 | M2/549 | M3/129 | XV/23 | |
| M2/8 | A | NA | NA | NA | NA | - | - | - | - | - |
| M2/355 | A | NA | NA | NA | NA | A | - | - | - | - |
| M2/549 | A | NA | NA | NA | NA | A | A | - | - | - |
| M3/129 | A | NA | NA | NA | NA | A | A | A | - | - |
| XV/23 | NA | NA | NA | A | NA | NA | NA | NA | NA | - |
| M2/716 | NA | NA | NA | A | NA | NA | NA | NA | NA | A |
Discussion
All six ramosus mutants showed single gene, recessive inheritance and clear expression under an 18-h photoperiod. Four mutants (M2/8, M2/355, M2/549 and M3/129) proved to be allelic with rms1 and two (XV/23 and M2/176) were allelic with rms4. This brings the total of the known mutations to eleven for rms1 and five for rms4. Currently identified mutations at loci Rms1 through Rms5 and their suggested allele numbers are summarised in Table 3. We have used a revised symbol notation as described at the foot of Table 3.
While the new mutants provide no evidence of additional ramosus genes, they do add significantly to the array of mutations at known rms loci available for study at the physiological, biochemical and molecular level. In particular, the exploratory value of mutants is increased where we have several alleles in the same genetic background as is the case for rms1 alleles 6 (M2/8), 7 (M2/355) and 8 (M2/549) in the Borek background. Again, exploration of double mutants is best done using isogenic lines and we now have one rms4 mutant (M2/716) and three rms1 mutants in a Borek background to pursue this goal. Based on F2 data from crosses between Borek and its mutant lines, the three mutations rms1-6, rms1-7 and rms1-8 appear similar in the severity of their phenotypic effects. The ratio TLL:TL (see Fig. 1) ranged from 1.26 ± 0.13 for rms1-6 to 1.32 ± 0.13 for rms1-8 with the value for rms1-7 segregants intermediate at 1.30 ± 0.20 (all differences not significant, P>0.7). Hence all three mutations caused a similar reduction in apical dominance based on this index of branching intensity. Compared with WT F2 segregants, rms1-6, rms1-7 and rms1-8 F2 plants showed a 16, 14 and 10% reduction in stem length between nodes 1 and 9, and a 22, 15 and 25% reduction in total length of the main stem, respectively. These observations are in accordance with other reports (2, 7, 15) that mutations in genes Rms1 through Rms5 generally cause a reduction in internode length and plant height.
A note of caution needs be raised regarding the mutant M3/129 and its initial line 9-76. This initial line was atypical of a WT line as it displayed a weak tendency to form basal branches under an 18-h photoperiod in a manner reminiscent of the mutant line K586 (see 2). Thus, while the difference between M3/129 and line 9-76 is clear and undoubtedly due to a mutation in the Rms1 gene, we can not exclude the possibility that the initial line may itself possess some weakly expressed mutation in an Rms gene. Alternatively, the quantitative genetic background in line 9-76 may simply confer an increased tendency for basal branching.
Acknowledgments. We thank Drs N. Naidenova and M. Vassileva for providing seed of the six mutants and their initial lines, and the Australian Research Council for financial support.
Table 3. Details, including suggested allele numbers, for 26 ramosus mutants which have now been assigned to one of the loci Rms1 through Rms5 as a result of allelism tests.
| Mutant
allele* |
Mutant
line |
Initial
line |
Mutagenic
agent |
Author
of
mutant line |
Key
genetic
references
|
| rms1-1 | WL5237 | Parvus | X-rays | S. Blixt |
7 |
| 2 | WL5147 | Weitor | X-rays | S. Blixt |
2 |
| 3 | WL5918 | Raman | Gamma-rays | M. Vassileva |
2 |
| 4 | Wt15236 | Paloma | Fast-n+NEU | W.K. Swiecicki |
2 |
| 5 | Wt15240 | Kaliski | NEU | W.K. Swiecicki |
2 |
| 6 | M2/8 | Borek | Gamma-rays | N. Naidenova |
16 |
| 7 | M2/355 | Borek | Gamma-rays | N. Naidenova |
16 |
| 8 | M3/549 | Borek | Gamma-rays | N. Naidenova |
16 |
| 9 | M3/129 | 9-76 | Gamma-rays | N. Naidenova |
16 |
| 10 | M3T-884 | Tйrиse | EMS | C. Rameau |
15 |
| 11 | M3T-988 | Tйrиse | EMS | C. Rameau |
15 |
| rms2-1 | K524 | Torsdag | EMS | K. Sidorova |
2 |
| 2 | WL5951 | Parvus | EMS | S. Blixt |
2 |
| rms3-1 | K487 | Torsdag | NMU | K. Sidorova |
2 |
| 2 | K564 | Torsdag | EMS | K. Sidorova |
2 |
| 3 | WL6042 | Meteor | Gamma+EMS | M. Vassileva |
2 |
| 4 | T2-30 | Tйrиse | EMS | C. Rameau |
15 |
| 5 | M2T-32 | Tйrиse | EMS | C. Rameau |
15 |
| rms4-1 | K164 | Torsdag | EMS | K. Sidorova |
2 |
| 2 | Wt15242 | Paloma | NEU | W.K. Swiecicki |
2 |
| 3 | M3T-946 | Tйrиse | EMS | C. Rameau |
15 |
| 4 | M2/716 | Borek | Gamma-rays | N. Naidenova |
16 |
| 5 | XV/23 | Aurelia | Fast-neutrons | M. Vassileva |
16 |
| rms5-1 | Wt15244 | Porta | Fast-neutrons | W.K. Swiecicki |
1,2 |
| 2 | Wt10852 | Paloma | NEU | W.K. Swiecicki |
2 |
| 3 | Wt15241 | Paloma | NEU | W.K. Swiecicki |
2 |
* We have used here a revised symbol notation. For example, the gene written by Arumingtyas et al. (2) as Rms-1 is now written Rms1. Each mutant allele of Rms1 is identified by a number separated from the base symbol by a hyphen, e.g. rms1-1, rms1-2, etc.
______________________
1. Apisitwanich, S., Swiecicki, W.K. and Wolko, B. 1992. Pisum Genetics 24:14-15.
2. Arumingtyas, E.L., Floyd, R.S., Gregory, M.J. and Murfet, I.C. 1992. Pisum Genetics 24:17-31.
3. Beveridge, C.A., Murfet, I.C., Kerhoas, L., Sotta, B., Miginiac, E. and Rameau, C. 1997. Plant J. 11:339-345.
4. Beveridge, C.A., Ross, J.J. and Murfet, I.C. 1994. Plant Physiol. 104:953-959.
5. Beveridge, C.A., Ross, J.J. and Murfet, I.C. 1996. Plant Physiol. 110:859-865.
6. Blixt, S. 1968. Agri Hort. Genet. 26:136-148.
7. Blixt, S. 1976. Agri Hort. Genet. 34:83-87.
8. Doroshenko, A.V. and Rasumov, V.I. 1929. Trudy prikl. Bot. Genet. Selek. 22:219-276.
9. Floyd, R.S. and Murfet, I.C. 1986. Pisum Newsletter 18:12-15.
10. Lamprecht, H. 1950. Agri Hort. Genet. 8:1-6.
11. Monti, L.M. 1970. Pisum Newsletter 2:21-22.
12. Monti, L.M. and Scarascia-Mugnozza, G.T. 1967. Genetica Agraria 21:301-312.
13. Murfet, I.C. and Reid, J.B. 1985. In
The Pea Crop : a Basis for Improvement,
Eds P.D. Hebblethwaite, M.C. Heath and T.C.K. Dawkins, Butterworths,
London, pp 67-80.
14. Murfet, I.C. and Reid, J.B. 1993. In Peas
- Genetics, Molecular Biology and Biotechnology,
Eds R. Casey and D.R. Davies, CAB International, Wallingford, U.K.,
pp 165-216.
15. Rameau, C., Bodelin, C., Cadier, D., Grandjean, O., Miard, F. and Murfet, I.C. 1997. Pisum Genetics. 29:
16. Symons, G.M. and Murfet, I.C. 1997. Pisum Genetics. 29: