|
Pisum Genetics |
Volume 24 |
1992 |
Research Reports |
pages 86-89 |
Internode length in Pisum. Two further lv mutants.
|
Weller, J.L., Murfet, I.C. and Reid, J.B. |
Department of Plant Science University of Tasmania Hobart, Tasmania 7001, Australia |
The mutant alleles of four major internode length genes in pea, La, Cry, Sln and Lv, have now been shown to result in a phenotype with elongated internodes. The double mutant la crys has long, thin internodes and a phenotype known as slender (1, 2). The la crys slender plants behave as if saturated with gibberellins (GAs), and the slender phenotype is expressed even in genotypes grossly deficient in GAs (6). More recently, a new mutant with elongated internodes and a phenotype similar to la crys slender plants, at least in regard to the lower internodes, was shown to result from the sln mutation (9). Plants homozygous for sln have elevated levels of C19 gibberellins. It is suggested (9) that the sln mutation may block catabolism of GA20 in the seeds and that this results in elevated levels of GA1 in the young seedling.
Plants homozygous for the mutant allele lv have elongated internodes, a semi-etiolated appearance, and an enhanced response to applied GA1 (5, 8). The lv allele is thought to block the action of light-stable phytochrome (5), and differences between Lv and lv plants are maximised under a regime of continuous light from cool white fluorescent tubes (10) since this light source provides only a negligible level of far-red light.
We report here on inheritance and allelism tests with two further pea mutants, L80m and Wt10895, with elongated internodes. Both mutants showed monogenic recessive inheritance and proved to be allelic with lv.
Materials and Methods
The elongated mutant L80m arose spontaneously in our stocks of line L80 (Lamm L30). The elongated mutant Wt10895 was obtained at Wiatrowo Plant Breeding Institute by Prof Dr W.K. Swiecicki following treatment of the dwarf cultivar Paloma (Wt3527) with 200 r Nf (fast neutrons)/0.014% NEU (nitroso ethyl urea). These two elongated mutants were tested for allelism with lv by crossing with NEU3 (lv; 8) an elongated mutant obtained by Dr T.A. LaRue from cv. Sparkle. Hobart lines L2 (Lamm line 2) and L85 were also used in crosses. Lines 2 (le la Cry Lv; 2, 4), 80 (le La Cry Lv; 3), 85 (le La cryc Lv; 4) and Sparkle (le La cryc Lv; 7) are all phenotypically dwarf.
The plants were grown in 14 cm slim line pots in a 1:1 (v:v) mixture of vermiculite and 10 mm dolerite chips topped with 4 cm of sterilised peat-sand potting mixture. Nutrient was supplied once a week in the form of Total Growth Nutrient (R and D Aquaponics, Sydney). The measurements in Figs 1, 2 and 4 were obtained from plants grown in growth cabinets under continuous light from cool white fluorescent tubes (Figs 1 and 4, 90 mmol m-2 s-1 and Fig. 2, 6 mmol m-2s-1) at a constant temperature (Figs 1 and 4, 20°C, and Fig. 2, 17.5°C). These conditions were used during the early stages of growth to screen the genotypes. After separation was effected (usually 2-3 weeks), the plants were transferred to normal glasshouse conditions for growth to maturity. The data in Fig. 3 come from plants grown under an 8 h photoperiod (8 h daylight, 16 h dark; temperature 18-22/16°C). Node counts commenced from the first scale leaf as node 1.
Results and Discussion
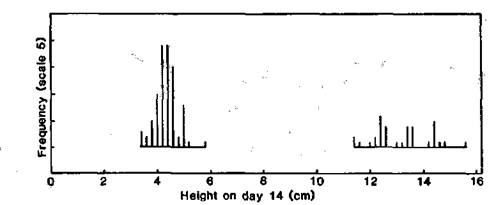
The data in Fig. 1 were obtained from several progenies segregating L80-type (dwarf) and L80m-type (elongated) plants. The observed numbers of 85 dwarf and 35 elongated segregates are in good accordance with a 3:1 ratio (c2 = 1.11, P > 0.2) and indicate monogenic recessive inheritance of the mutant type.
Fig. 1. Distribution of height (soil surface to shoot apex) at day 14 for line 80 progenies segregating for dwarf (L80) and elongated (L80m) types. Conditions: continuous white fluorescent light (90 mmol m-2 s-1) at 20°C.
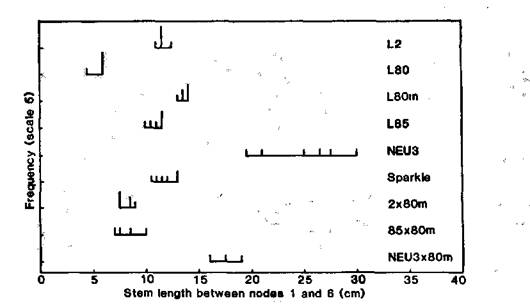
Fig. 2. Distribution of stem length between nodes 1 and 6 for dwarf lines L2, L80, L85 and Sparkle, elongated mutant lines L80m and NEU3, and F1 plants of crosses 2 x 80m, 85 x 80m and NEU3 x 80m. Conditions: continuous white fluorescent light (6 mmol m-2 s-1) at 17.5°C.
Elongated segregates bred true in subsequent generations. The mutant-type segregates were also readily distinguished under this fluorescent light regime by reason of the epinasty of the leaf petioles, and the failure of the leaflets to expand normally. The data in Fig. 2 indicate mutant L80m is allelic with lv since the F1 of cross L80m x NEU3 (lv) was also an elongated type with a length intermediate between that of the two parents. The F1 plants of crosses L80m (le La Cry lv80m) x L2 (le la Cry Lv), and L80m x L85 (le La cryc Lv) were dwarf and somewhat shorter than either parent, presumably because the F1 plants possess all three dominant alleles La, Cry and Lv (see 3).
The F2 data from cross Wt3527 (dwarf initial line) x Wt10895 (elongated mutant) likewise indicate monogenic recessive inheritance of the mutant type (Fig. 3). The observed numbers of 43 dwarf and 10 elongated segregates are in good accordance with a 3:1 ratio (c2 = 1.06, P > 0.3). The elongated types bred true in subsequent generations. The data from the cross Wt10895 x NEU3 (lv) indicate that mutant Wt10895 is also allelic with lv, since the F1 plants were elongated types intermediate in length between the two parents (Fig. 4). In contrast, the F1 of crosses NEU3 x Wt3527 and Wt10895 x Sparkle were dwarf in phenotype and no greater in length than cv. Sparkle (Fig. 4).
Four elongated mutants have now been identified as resulting from mutation of gene Lv (8; Figs 2 and 4). The relative strengths of the four alleles lvNEU3 (type line NEU3), lvR83 (type line R83), lv80m (type line L80m) and lv10895 (type line Wt10895) remain to be determined.
Acknowledgements. We thank Dr T.A. LaRue and Professor Dr W.K. Swiecicki, and the late Professors R. Lamm and G.A. Marx, for providing material, Peter Bobbi and Katherine McPherson for technical assistance, and the Australian Research Council for financial support.
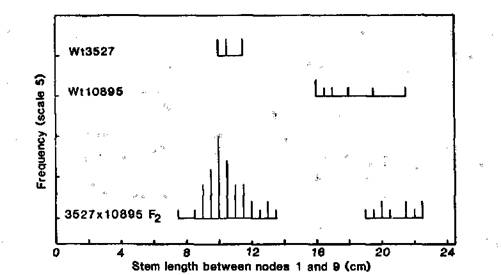
Fig. 3. Distribution of stem length between nodes 1 and 9 for the elongated mutant line Wt10895, the initial line Wt3527 (cv. Paloma), and the F2 of cross 3527 x 10895. Conditions: photoperiod 8 h (8 h daylight at about 18-22°C and 16 h dark at 16°C).
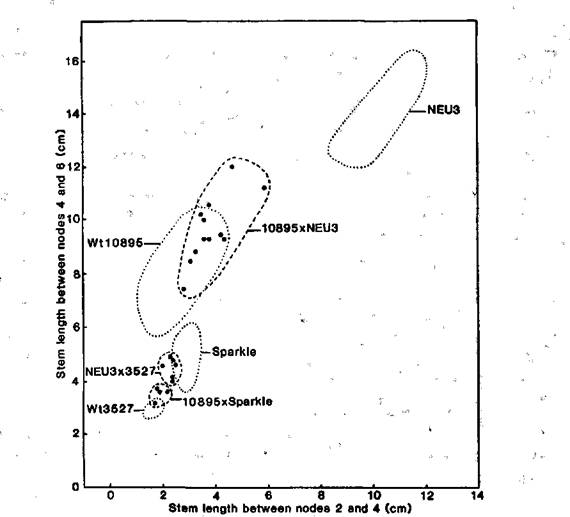
Fig. 4. Stem length between nodes 4 and 6 plotted against stem length between nodes 2 and 4 for mutant line NEU3 and its initial line Sparkle, mutant line Wt10895 and its initial line Wt3527, and the F1 (●) of crosses 10895 x NEU3, NEU3 x 3527 and 10895 x Sparkle. The plots for the parental lines fell within the respective dotted boundaries (n = 6 except for Wt10895 where n = 18). Conditions: continuous white fluorescent light (90 mmol m-2 s-1) at 20°C.
deHaan, H. 1927. Genetica 9:481-497.
Lamm, R. 1947. Hereditas 33:403-419.
Marx, G.A. 1974. Pisum Newsl. 6:32-34.
Murfet, I.C. 1988. Ann. Bot. 61:331-345.
Nagatani, A., Reid, J.B., Ross, J.J., Dunnewijk, A. and Furuya, M. 1990. J. Plant Physiol. 135:667-674.
Potts, W.C., Reid, J.B. and Murfet, I.C. 1985. Physiol. Plant. 63:357-364.
Reid, J.B. 1987. Pisum Newsl. 19:52-54.
Reid, J.B. and Ross, J.J. 1988. Physiol. Plant. 72:595-604.
Reid, J.B., Ross, J.J. and Swain, S.M. 1992. Planta 188:462-467.
Weller, J.L. and Reid, J.B. 1993. Planta 189:15-23.