THE PHYSIOLOGY OF A WILTY MUTANT OF PISUM SATIVUM UNDER SIMULATED WATER
STRESS CONDITIONS
Donkin, M. E., T. Hull, and E. S. Martin
Rumleigh Experimental Station, Plymouth, U. K.
Wang, T. John Innea Institute, Norwich, U. K.
Modifications of plant funotion caused by a single gene mutation
can reveal insights into fundamental aspects of the physiology of
plants, particularly in relation to their environment. Nowhere is this
more apparent than in the water relations of crops. Mutations causing
wilting, for example, have been reported in tomato, potato, corn and
peppers. The cause of the abnormality has been ascribed to alterations
in hormone balance, blocked vessel elements and changes in the ionic
balance in the guard cells of the plants.
In 1976 Marx (2) described a wilty mutant of pea which wilted under
conditions of water stress. We have recently carried out studies on the
water relations and stomatal behavior of this mutant (called "Wilty" or
JI 1069) to compare them with other non-wilty lines (JI's 1180, 1194 and
74) of similar phenotype (1). "Wilty" leaves had a lower percentage
water content, a lower water potential and a lower diffusive resistance.
In addition the guard cells of "Wilty" were slighty larger than those of
several other non-wilty lines.
When stomatal behavior of "Wilty" was investigated it was found
that the aperture of stomata on epidermal samples taken from plants in
the light and dark period of a diurnal rhythm were larger for the
"Wilty" compared to a non-wilty line (JI 1180) . However, stomatal
ones on detached epidermis to light, CO2- and KCl concentration (in
the medium) were similar in "Wilty" and non-wilty lines. There was also
no difference in the response of stomata to the hormone, abscisic acid
(ABA).
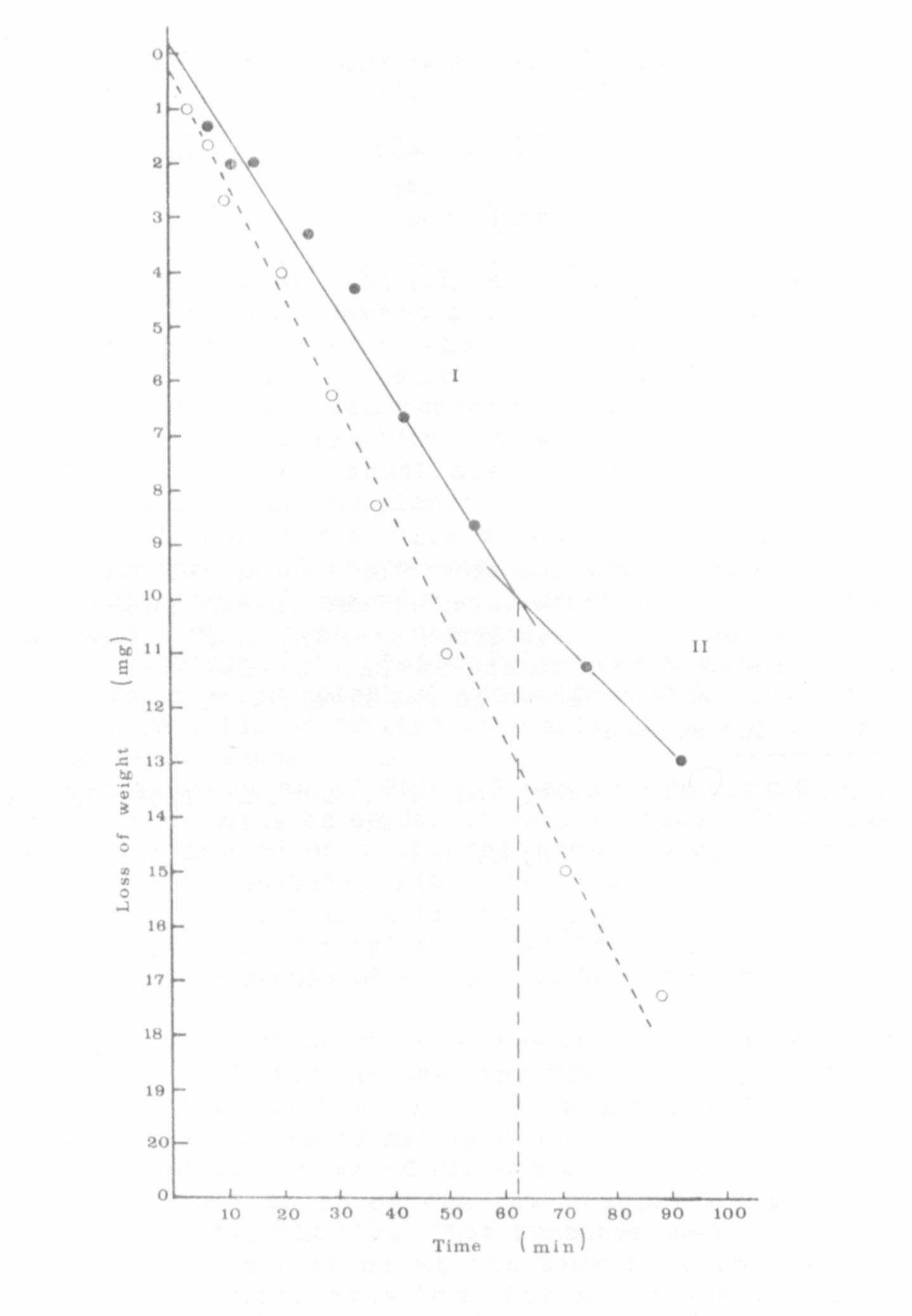
Preliminary studies of water loss from detached leaflets (to simu-
late water stress) showed (Fig. 1) that the "Wilty" line tends to lose
water at a rate similar to that of a non-wilty pea during the first
phase (I) of water loss, which is mainly due to stomatal and cuticular
transpiration. However, after 60 min the water loss of the non-wilty
pea begins to slow down as it enters the transition phase of the
response curve (Phase II, Fig. 1). This response does not occur in the
"Wilty" since water loss continues at the same rate for up to 90 min
after excision. This indicates that the stomata fail to close as
quickly in "Wilty" when it is under water stress. At the end of the ex-
periment atrip* of upper epidermis were removed from each leaflet of the
"Wilty" and non-wilty samples and stomatal apertures were measured under
the microscope; Table 1 shows that the stomata of "Wilty" leaflets were
slightly more open than those of the non-wilty plant.